
Скорость бега от чего зависит: От чего зависит скорость бега и как ее увеличить?
от чего зависит, техника — Рамблер/спорт
Составляющие скорости бегуна Самый быстрый бег напрямую зависит от длины бегового шага, который обязан быть оптимальным. и обеспечивать спортсмену мимолётное расслабление мышечных групп ног и помогает беречь силы. А интенсивность в свою очередь — от отталкивания от поверхности. Поэтому дабы увеличить скорость, на старт тратим минимум времени, так при его сокращении, фаза переноса маховой нижней конечности тоже сократится. Нельзя ждать соприкосновения конечностей с опорой, а стараться активно уменьшить момент полёта, ведь стремительность зависит и от частоты совершаемых шагов. Помимо этого, лишний вес и вредные привычки в таком деле играют далеко не последнюю роль.
Блок похожие статьи
Техника быстрого бега Дабы увеличить скорость до максимально возможной, нужно обязательно следовать данной технике:
Руки под 90-градусным углом согнуть в локтях. Необходимо осуществлять движения вперёд — назад в темпе с ногами, это способствует повышению аэродинамики. Амплитуда верхних конечностей также задает интенсивность. Чтобы от дорожки оттолкнуться с силой, ноги должны выполнять мягкие и пружинистые действия и начинать движения мгновенно. При этом и большую роль играет соблюдение правильного наклона кора. Для развития высокого темпа его нужно слегка наклонить вперёд. Иначе сильное склонение будет способствовать замедлению работы ног. Также уклон зависит от физической подготовки и особенностей телосложения человека.
Амплитуда верхних конечностей также задает интенсивность. Чтобы от дорожки оттолкнуться с силой, ноги должны выполнять мягкие и пружинистые действия и начинать движения мгновенно. При этом и большую роль играет соблюдение правильного наклона кора. Для развития высокого темпа его нужно слегка наклонить вперёд. Иначе сильное склонение будет способствовать замедлению работы ног. Также уклон зависит от физической подготовки и особенностей телосложения человека.
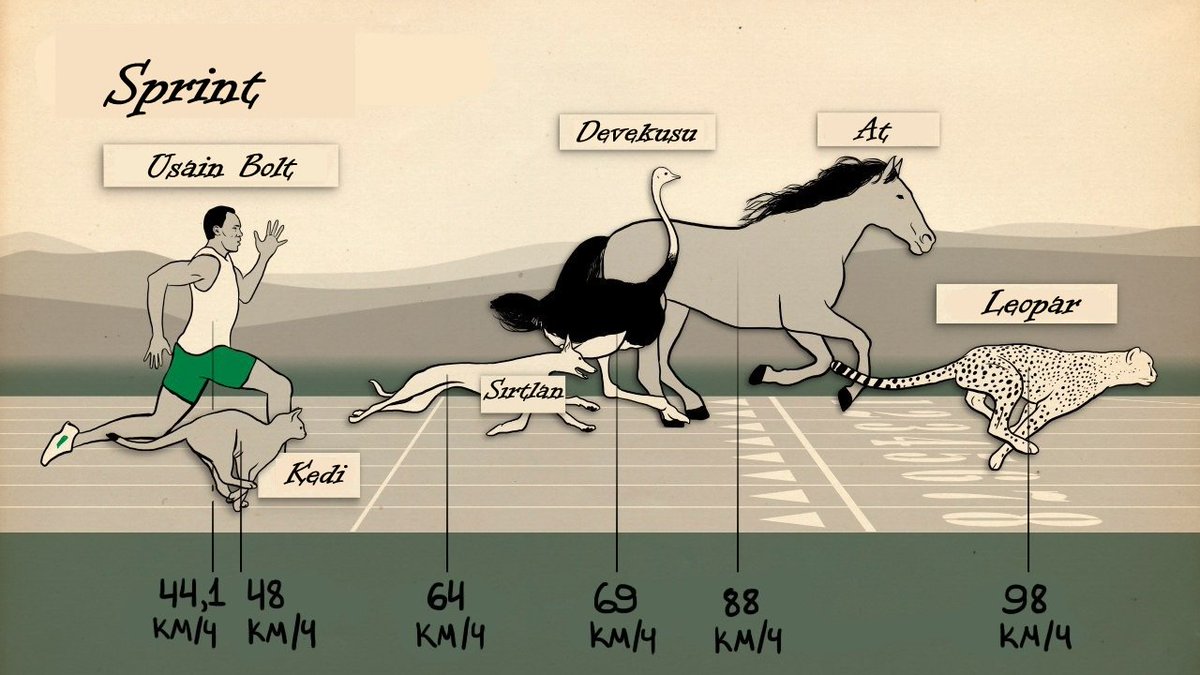
Быстрый бег Считается, что быстрый бег рассчитан на профессиональных легкоатлетов, так как это тренировка или кросс. Но также скорый бег — достаточно широкое понятие. Ведь темп в три минуты на один км. для соревнования довольно быстрый, а для длинного марафона – медленный. В таких забегах средняя скорость должна составлять от 22 км/ч, а максимальная — 43 км в час, этот рекорд скорости бега поставил Усейн Болт. Его показатели выше на 21 километр в час, чем у других марафонцев. Исследования показали, что люди могут развить до 63 км/ч. Придерживаясь данной технике по увеличению скорости, можно добиться хороших результатов. Не маловажно тренироваться и упорно работать над своим телом. Читать далее
Придерживаясь данной технике по увеличению скорости, можно добиться хороших результатов. Не маловажно тренироваться и упорно работать над своим телом. Читать далее
Другие материалы по теме:
Развитие общей выносливости и бег: какова их взаимосвязь
Основные методы бега
Эффективный комплекс бега для правильной техники пробежек
От чего зависит ваш естественный темп бега
У каждого бегуна есть свой естественный темп. Вы автоматически поддерживаете этот темп, когда собираетесь размеренно и спокойно пробежать заданную дистанцию, например 8 километров, или бежать в течение определённого времени, например 45 минут.
Естественный темп меняется в зависимости от физической формы и даже от самочувствия в конкретный день.
Что определяет естественный темп бегуна? Учёные, исследующие физические упражнения, мало интересовались этим вопросом, поэтому ответ найти не так просто. Однако есть несколько исследований, которые хоть немного проясняют ситуацию.
Естественный темп и молочная кислота
KROCKY MESHKIN/Flickr. com
comВ исследовании 2001 года учёные из Университета Удины в Италии проверяли гипотезу, согласно которой естественный темп бегуна зависит от уровня молочной кислоты в крови.
Они предполагали, что на естественный темп в первую очередь влияет максимальное устойчивое состояние по лактату крови. То есть это самый быстрый темп, который способен поддерживать бегун без накопления молочной кислоты до концентрации, при которой чувствуется усталость.
В исследовании участвовали восемь бегунов-любителей. Для начала определили их лактатный порог — максимальную скорость и частоту сердечных сокращений до повышения уровня молочной кислоты в крови. Затем участников попросили бегать в естественном темпе в течение часа.
В среднем все бегуны закончили часовую пробежку с максимальным устойчивым состоянием по лактату.
Максимальное устойчивое состояние по лактату — это интенсивность физической нагрузки, при которой создаётся максимально устойчивое состояние между выходом в кровь молочной кислоты и её утилизацией.

И несмотря на то, что скорость бега для достижения максимального устойчивого состояния по лактату у всех участников сильно отличалась (одни бежали быстрее, чем другие), различия в естественном темпе на протяжении часа у всех бегунов были минимальны.
Это привело учёных к мысли, что, помимо необходимости избегать накопления молочной кислоты в крови, есть и другие факторы, влияющие на естественную скорость бега.
В исследовании учёных из Университета Удины было две проблемы. Во-первых, нет доказательств, что темп бега строго лимитируется уровнем лактата в крови. В коротких гонках, например, бегуны часто достигают уровня молочной кислоты, превышающего лактатный порог.
Если такая высокая концентрация молочной кислоты допустима в коротких гонках, почему в других обстоятельствах должны действовать жёсткие ограничения по уровню лактата?
Вторая проблема гипотезы о молочной кислоте заключается в том, что в человеческом теле нет механизма, с помощью которого уровень молочной кислоты мог бы повлиять на темп бега, даже если он вызывает усталость в мышцах.
Если уровень лактата в крови регулирует темп бега с помощью усталости, тогда каждый спортсмен всегда бежал бы в одном темпе, при котором поддерживается нужный уровень молочной кислоты. А как мы знаем, естественный темп бега может меняться даже от настроения.
А вот что исследователи из Университета Удины упустили, так это роль мозга в регулировании физической активности. Ведь именно мозг сообщает мышцам, насколько сильно они должны напрягаться при выполнении любых упражнений.
Поэтому настоящее объяснение феномена естественного темпа, должно быть, кроется в мозгу.
Естественный темп определяют два желания
Эта истина была обнаружена в другом исследовании 2001 года, проведённом учёными из Университета Уэйна в Небраске.
Восемнадцать мужчин и женщин попросили тренироваться в течение 20 минут, выбрав подходящую для них интенсивность. Всего было три упражнения: тренировка на беговой дорожке, занятие на велотренажёре и подъём по лестнице на степпере.
По итогам эксперимента физиологические показатели после каждого упражнения были разными. В среднем после упражнений на велотренажёре у участников отметили более высокий VO2maх, чем после тренировок на степпере и беговой дорожке, а после степпера у них был более высокий пульс, чем после педалирования и бега.
В среднем после упражнений на велотренажёре у участников отметили более высокий VO2maх, чем после тренировок на степпере и беговой дорожке, а после степпера у них был более высокий пульс, чем после педалирования и бега.
Как бы то ни было, во время всех трёх упражнений они чувствовали себя примерно одинаково. Из этого можно сделать вывод, что естественный темп бега и предпочитаемая интенсивность выполнения других упражнений не полностью определяются физиологией: большое значение также имеют ощущения. А где они рождаются? В мозгу.
Другие исследования показали сходные результаты. Когда участникам разрешали выбирать интенсивность, они всегда тренировались на границе комфортной зоны с оглядкой на продолжительность тренировки. Почему именно этот уровень?
Возможно, это некий компромисс между двумя желаниями, которые возникают в мозгу во время тренировки: как можно быстрее выполнить задание и чувствовать себя комфортно.
Итак, ваш естественный темп бега, будь то 7, 6 или даже 5 минут на километр, показывает ваш внутренний компромисс и индивидуальные беговые способности.
Но как ваш естественный темп влияет на достижение целей? Помогает ли он повысить уровень беговой подготовки или, наоборот, препятствует этому?
Стоит ли всегда бегать в естественном темпе
Естественный темп бега приблизительно совпадает с интенсивностью, при которой вы остаётесь в жиросжигательной зоне ЧСС. Так что этот темп идеален для долгих пробежек с целью сжигания жира и тренировки выносливости.
Поскольку естественный темп не требует таких энергетических затрат, как быстрый, с его помощью вы можете постепенно увеличить пробегаемое расстояние, а также развить экономичность бега. Так что бег в естественном темпе стоит включить в свой план тренировок.
Однако некоторые бегуны слишком часто, а то и постоянно бегают в естественном темпе. Как минимум раз в неделю стоит включать в свои тренировки бег с высокой интенсивностью. Он улучшит вашу физическую форму и дополнит преимущества бега в естественном темпе.
Не обязательно устраивать изнурительные интервальные сессии и бег за лактатным порогом. Вы можете просто немного разнообразить тренировки, сочетая бег в естественном темпе и небольшое количество ускорений.
Вы можете просто немного разнообразить тренировки, сочетая бег в естественном темпе и небольшое количество ускорений.
Естественный темп + ускорение
Хороший пример — тренировка фартлек, в которой вы чередуете короткие спринты (например, шесть раз по 45 секунд на протяжении 5 километров) с размеренным бегом, что позволяет сделать тренировку легче и в то же время немного развить скорость и сопротивление усталости.
Пробежки по методике фартлек отлично подходят для начального уровня подготовки, когда вы ещё не готовы к изматывающим интервальным тренировкам.
Ещё один вариант — прогрессивный бег. Это спокойный, размеренный бег (обычно от 2 до 5 километров в марафонском или полумарафонском темпе) с ускорением в конце.
Такой метод тренировок тоже хорошо работает на базовом этапе, если вы ещё не готовы к интервальным занятиям, а также в любой момент тренировок, когда вы запланировали «лёгкий день», но чувствуете себя отлично и хотите немного увеличить нагрузку.
И наконец, прогрессивный бег подойдёт для перехода от длительных пробежек к пиковой фазе тренировок, когда вы хотите превратить общую выносливость, развитую в процессе долгих пробежек, в выносливость для соревнований.
Кроме понимания, когда стоит использовать естественный темп бега, также полезно просто понаблюдать за ним. Замечать изменения в вашем естественном темпе — это самый простой и вместе с тем мотивирующий способ отслеживать ваш уровень подготовки.
Чем больше ваш прогресс, тем быстрее естественный темп. Вы будете бежать всё быстрее и быстрее, при этом сохраняя ощущение комфорта. Просто доверьтесь своим чувствам и наслаждайтесь результатами.
Как увеличить скорость бега
Об этой теме написано уже немало, но давайте еще раз разберемся в тонкостях.
Спортсменам приходится преодолевать собственные возможности, чтобы выйти на лучшие показатели. И профессиональные легкоатлеты, и просто любители бега, стремясь к совершенству, хотят увеличить свою скорость бега. Бежать быстро и долго — мечта всех бегунов без исключения! Бесконечно увеличивать скорость невозможно, у каждого есть свой предел физических способностей, все-таки генетика делает свое дело, но добиться максимума можно систематически тренируясь и поставив перед собой мощную мотивацию.
Факторы, влияющие на скорость бега
На беговую скорость влияют врожденные факторы, которые нельзя изменить кардинально. От генетики не убежать. Только регулярные тренировки с хорошей самоотдачей и преодолением собственных показателей способны максимально раскрыть потенциал спортсмена.
Физиологически, скорость бега зависит от процессов, происходящих в нервных тканях, что отражается на частоте сокращений мышц, способности быстрого расслабления и способности ускоряться.
Кроме врожденных факторов на спортивные результаты могут влиять:
- Физическое и моральное состояние.
- Переутомление как физическое, так и нервное.
- Состояние психики.
- Погодные условия и экипировка.


Составляющие скорости бега — это длина шага и его частота. Именно на это можно обратить внимание во время тренировок.
Особенности тренировок
Результат по улучшению скорости бега не заставит себя ждать, если будут соблюдены некоторые простые условия:
- Есть конкретная цель (преодолеть дистанцию за определенное время).
- Регулярные тренировки с полной отдачей возможностей.
- Есть грамотная программа тренировок.
Мышцы ног имеют решающее значение для скорости бега. Поэтому всегда необходимо включать упражнения на силу ног в программу занятий.
Второй, не менее важный момент — прыгучесть, умение спортсмена отталкиваться и в прыжке мгновенно разгоняться. Следовательно, надо выполнять не только статические упражнения, но и динамические. Чтобы почувствовать скорость в ногах, необходимо выполнять ускорения, например, на 60 — 150 м, где надо ускоряться сразу и до финиша.
Многие тренера советуют не забывать включать в свои тренировки работу над скоростными показателями, прыжки и силовые упражнения, а также отрабатывать технику бега, выполняя СБУ.
Упражнения на развитие скорости бега
Данные упражнения можно включать в программу занятий «силового» дня.
Полуприсед в прыжке. 10 повторений на прыжки из положения полуприсед на максимальную высоту.
Выпад в прыжке. 10 повторений прыжков из положения выпад со сменой ног в момент прыжка.
Прыжки через барьер. На беговой дорожке расставляются 10 одинаковых барьеров в шахматном расположении на расстоянии 60 см. Через препятствия важно перескакивать.
Прыжки. 10 прыжков на одной ноге с полной отдачей и попыткой зависания в воздухе.
Среди других упражнением можно выполнять занятия со скакалкой, пробежку с подскоками (ноги растягивать в высоту и длину). Отличную силовую нагрузку дает работа в паре двух спортсменов на сопротивление.
Интервальные тренировки следует проводить в день, когда нет тяжелых силовых упражнений.
Подсчет своей скорости бега (темп и скорость)
Подготовка к преодолению дистанции за определенное время заключается в правильном контроле темпа движения, чтобы достичь желаемого результата.
Лучшим считает равномерное преодоление дистанции. Для этого следует поделить всю дистанцию на отдельные равные участки и представлять, с какой скоростью надо преодолевать очередной отрезок, то есть сделать раскладку по темпу.
Бег на полумарафонскую дистанцию требует деление на 1 км и каждые 5 км. Если планируется преодолеть полумарафон за полтора часа, то время на километр отводится 4 мин 30 сек, а на 5 км — 21 мин и 40 сек.
Еще одна важная составляющая — скорость. Здесь надо учитывать длину дистанции и тренировки проводить с запасом времени и скорости. Принцип достаточно простой, но высчитывать постоянно необходимые показатели не имеет смысла. Для этого можно составить специальную таблицу уже содержащую все данные на основные дистанции или воспользоваться калькулятором темпа и скорости. Такие калькуляторы в большом количестве вариантов представлены в интернете. Достаточно ввести необходимые данные, чтобы получить требуемые расчеты.
Здесь надо учитывать длину дистанции и тренировки проводить с запасом времени и скорости. Принцип достаточно простой, но высчитывать постоянно необходимые показатели не имеет смысла. Для этого можно составить специальную таблицу уже содержащую все данные на основные дистанции или воспользоваться калькулятором темпа и скорости. Такие калькуляторы в большом количестве вариантов представлены в интернете. Достаточно ввести необходимые данные, чтобы получить требуемые расчеты.
Таблица расчета темпа и скорости
Таблица расчета темпа и скорости для разных дистанций, в том числе для полумарафона и марафона
Во время тренировок многие спортсмены используют наручный секундомер с пульсометром, который помогает рассчитать нагрузку, темп и скорость для данной дистанции.
Советы по увеличению скорости бега
Выносливость и способность поддерживать необходимый темп вырабатывается постоянными тренировками с полной самоотдачей.- Тренироваться надо начинать в обычном для себя темпе с постепенным наращиванием его.
- Марафоны или полумарафоны надо бежать, не обращая внимания на других атлетов. Попытки догнать или опередить приведут к сходу с дистанции.
- Во время бега не надо контролировать технику бега. Это сбивает с ритма и снижает скорость. Техника должна быть отработана на тренировках.
- Искусственное увеличение шага может привести к травме связок.
- Тренировки можно проводить как утром, так и вечером.
- Пропуск тренировок приводит к нарушению режима. Заниматься и выдерживать график обязательно! Тем не менее, лучше отложить занятия если спортсмен недавно болел или переживал эмоциональные, нервные, умственные перегрузки.
- Если нет желания идти на тренировку, это может быть не проявлением расслабленности и лени, а последствия болезни или переутомления/перетренированностью. Это надо уметь различать.

Российские рекордсмены в беге на короткие дистанции
Бреднев Александр — один из самых именитых российских легкоатлетов, благодаря высочайшим показателям в беге. В чемпионате России Бреднев показал 10.38 сек. Спортсмен имеет чемпионский титул по бегу в помещениях на 60 метров.
В чемпионате России Бреднев показал 10.38 сек. Спортсмен имеет чемпионский титул по бегу в помещениях на 60 метров.
Мастеркова Светлана занесена в списки самых знаменитых легкоатлетов России. Начала бегать с 800 метров, была чемпионкой СССР перед его распадом. После перерыва на декретный отпуск и травмы ноги, Светлана вернулась в спорт при поддержке мужа и сразу взяла олимпийское золото в Атланте. Она четырехкратная чемпионка мира, чьи рекорды не смогли превзойти до сих пор.
Увеличение скорости бега. Советы для новичков и не только
Бег – это быстрое передвижение в пространстве с «фазой полёта», что позволяет спортсмену как значительно увеличивать скорость, так и переводить дыхание, беречь силы. Для того чтобы человек во время бега не сильно уставал и мог бежать дольше, мышцы его ног должны отдыхать.
• Все зависит от частоты.
Чем длиннее шаги при беге сделает человек, тем медленнее он будет бежать. Так происходит потому, что скорость бега существенно зависит от высокой частоты, а нет от максимальной длины шага. Для значительного увеличения скорости при беге нужно максимально сократить время отталкивания от беговой дорожки, и совершать движения с максимальной силой. В конечном итоге, длина шага не имеет очень большого значения для бегунов на скорость.
Так происходит потому, что скорость бега существенно зависит от высокой частоты, а нет от максимальной длины шага. Для значительного увеличения скорости при беге нужно максимально сократить время отталкивания от беговой дорожки, и совершать движения с максимальной силой. В конечном итоге, длина шага не имеет очень большого значения для бегунов на скорость.
• Нужна ли бегунам прыгучесть?
Помимо физических упражнений с дополнительными весами и тренировки выносливости, абсолютно любой спортсмен, который занимается бегом, должен уделять время прыжкам как в длину, так и в высоту. Поэтому прыгучесть занимает не последнее место для каждого легкоатлета.
• Отдых необходим во всем.
С одной стороны, для достижения результата важны повторы и постоянные тренировки, а с другой – просто необходимо отдыхать как на протяжении, так и после тренировки. Здоровый сон важен для абсолютно каждого спортсмена. Это способствует росту мышц и уменьшению количества травм.
• Советы для бегунов.
Для того чтобы значительно увеличить скорость при беге, необходимо сначала понять технику выполнения движений. По большей части, главным фактором является положение туловища бегуна. Для максимальной скорости, тело нужно сгибать вперёд и следить за осанкой. Но также надо постоянно помнить, что слишком сильный наклон делает спортсмена неустойчивым. И также немаловажным фактором является личная физическая подготовка и довольно хорошая генетика.
Существует масса споров на данную тему, и придумано множество методик тренировок. Так, многие спортсмены активно развивают особую разновидность выносливости – скоростную выносливость. Данный вид выносливости можно успешно тренировать при помощи, к примеру, «интервального бега». Суть такого типа бега, заключается в постоянном чередовании более быстрого и медленного темпа с небольшим отдыхом, так мышцы ног все время получают разные физические нагрузки. Тренеры разрабатывают все новые методики, но их суть, в большинстве своём, заключается в постоянных физических нагрузках на мышцы ног разной интенсивности.
А также довольно неплохим эффектом обладают упражнения которые используют методику, так называемого, темпового бега. Подразумевается, что спортсмен должен пробежать абсолютно всю заданную дистанцию как можно быстрее и техничнее. Огромное значение для значительного повышения скорости имеет способность выложить все силы на короткий отрезок дистанции, для преодоления дистанции как можно скорее.
• Заключение
Основной целью для абсолютно каждого спортсмена является как минимум – сохранить свой уровень и не ухудшить показатели, а как максимум – усовершенствование своих показателей. И теперь зная, как увеличить скорость вашего бега, все зависит только от вас, вашего стремления и моральной и физической подготовленности.
Техника бега при входе в поворот
Каждый бегун ставит перед собой определенные задачи. Чаще всего речь идет о повышении эффективности бега, и как следствие — улучшение беговых результатов. Чтобы этого достичь, нужно совершенствовать беговую технику. Одной из самых технически сложных траекторий бега являются повороты, на которых спортсмену приходится преодолевать давление центробежной силы. Именно от правильности входа в поворот зависит общая скорость преодоления дистанции, а значит, и конечный результат. И чем дистанция короче, тем техничнее должен быть бег. В этой статье поговорим об особенностях техники бега на поворотах, какое влияние они оказывают на скорость, рассмотрим основные ошибки, присущие неопытным бегунам, а также разберем упражнения, которые помогут усовершенствовать поворотную технику.
1. Где можно встретить поворотное движение?
Чаще всего повороты встречаются при беге по кругу на стадионе или в манеже. Но столкнуться с ними можно на любом другом участке дистанции, за исключением коротких отрезков по прямой.
Но столкнуться с ними можно на любом другом участке дистанции, за исключением коротких отрезков по прямой.
2. Особенности техники бега при входе в поворот
Техника бега при входе в поворот, или как ее еще называют «бег по поворотам (на виражах)», отличается от техники бега по прямой переходом на круговое движение. И чтобы эти изменения траектории существенно не повлияли на скорость, спортсмену следует придерживаться пяти основных правил:
ВАЖНО: Правила сформулированы, исходя из кругового движения против часовой стрелки, где поворотная сторона — левая, а противоположная повороту сторона — правая, что соответствует стандарту, установленному Международной ассоциацией легкоатлетических федераций.
- Правила постановки корпуса: приближаясь к центру поворота, слегка наклоните корпус влево для снижения давления центробежной силы. Чем больше радиус поворота, чем больше скорость, тем больше должен быть наклон.
- Правила движения рук: а) увеличьте скорость движения правой руки и снизьте скорость движения левой руки; б) вынесите правое плечо чуть вперед, отклонив левое плечо немного назад; в) локоть правой руки при движении назад отведите немного в сторону, а при движении вперед — больше внутрь; г) движение кисти левой руки должно идти больше наружу.
- Правила движения ног: а) немного сократите длину шага левой ноги; б) правую ногу ставьте на поверхность с небольшим разворотом стопы внутрь (то есть влево); в) уменьшите беговую посадку за счет снижения угла коленного сгиба и большего выпрямления ног. Это усилит нагрузку на колени, но позволит предотвратить резкое снижение скорости.
- Правила движения тела: все движения при входе в поворот и выходе из него осуществляйте плавно. В противном случае вас может вынести центробежной силой в сторону, в результате чего будут потеряны драгоценные секунды за счет преодоления лишних метров, а также есть риск столкновения с другими бегунами. Оптимально осуществлять наклон и выход из наклона в течение 20-30 м до и после поворота.
- Правило выхода из поворота: при выходе из поворота на прямую первые 20-30 м сохраняйте свободный размашистый бег и набранную скорость. Это называется «свободный ход» или бег по инерции, когда спортсмен не прилагает мышечных усилий для увеличения скорости. Это поможет избежать усиления мышечного перенапряжения, которое возникает при входе в поворот. Такой бег проводится на отрезке 8-10 м (4-5 беговых шагов). После «свободного хода» можно вновь включиться в активную работу и начать постепенно увеличивать скорость на прямой.

 Оптимально осуществлять наклон и выход из наклона в течение 20-30 м до и после поворота.
Оптимально осуществлять наклон и выход из наклона в течение 20-30 м до и после поворота. 3. Влияние поворотов на скорость
Бег на повороте менее эффективен и более энергозатратен, чем бег по прямой, из-за воздействия центробежной силы. Чтобы уменьшить ее влияние, придется отклониться от прямой постановки корпуса, что неизбежно приведет к снижению скорости движения за счет падения частоты, длины шагов и увеличения мышечной нагрузки. Это напрямую зависит от скорости бега, массы тела бегуна и радиуса поворота. Чем выше скорость и масса, чем меньше радиус — тем сильнее давление центробежной силы. В среднем разница в скорости на повороте и прямой на стадионе равна 0,2-0,3 с, а в манеже, где крутизна виража, как правило, больше — 0,5-0,8 с. Важность этих показателей зависит от длины пробегаемой дистанции. При беге на длинные дистанции угол виража меньше влияет на результат, чем на средних и особенно коротких дистанциях.
Это напрямую зависит от скорости бега, массы тела бегуна и радиуса поворота. Чем выше скорость и масса, чем меньше радиус — тем сильнее давление центробежной силы. В среднем разница в скорости на повороте и прямой на стадионе равна 0,2-0,3 с, а в манеже, где крутизна виража, как правило, больше — 0,5-0,8 с. Важность этих показателей зависит от длины пробегаемой дистанции. При беге на длинные дистанции угол виража меньше влияет на результат, чем на средних и особенно коротких дистанциях.
4. Оптимальное расположение на дистанции для входа в поворот
Чем выше радиус поворота, тем меньше физических и технических усилий требуется для его прохождения. Но чем дальше дорожка расположена от внутреннего края, тем длиннее пробегаемый круг. На стадионе длина каждой последующей дорожки больше на 7 м, в манеже — на 3 м. Поэтому оптимальные дорожки для поворота на стадионе — вторая (при четырех дорожках) и третья (при восьми-девяти дорожках), в манеже — вторая (при четырех дорожках) и третья (при шести дорожках).
5. Типичные ошибки в поворотной технике
Бегуны-любители иногда не проводят различий между бегом по прямой и повороту, совершая ряд ошибок, которые неизбежно сказываются на конечном результате. Среди них чаще всего встречаются следующие:
- Неправильный наклон корпуса за счет изгиба в пояснице и наклона головы на бок.
- Резкий наклон корпуса влево или резкий выход корпуса из наклона.
- Преждевременный наклон корпуса влево задолго до входа в поворот или затяжной выход корпуса из наклона после выхода на прямую.
- Чрезмерный наклон корпуса влево.
- Сильное сгибание ног в коленях.
- Одинаковая работа рук на протяжении всей дистанции.
6. Специальные упражнения для улучшения техники бега на повороте
Основное упражнение — короткое ускорение на повороте.
Тренировочная дистанция — 50-80 м. Начинать тренировки лучше с большего радиуса поворота на крайней дорожке стадиона или манежа, уменьшая радиус постепенно по мере улучшения техники бега.
Начинать тренировки лучше с большего радиуса поворота на крайней дорожке стадиона или манежа, уменьшая радиус постепенно по мере улучшения техники бега.
При выполнении упражнения движение начинается за 10-20 м от поворота. Скорость зависит от тренировочной дорожки — на крайней дороже она должна составлять 80-90% от максимальной, на 1-2 дорожках — 50-80% от максимальной.
Ускорение на повороте с выходом на прямую
Тренировочная дистанция — 80-100 м. Ускорения выполняются с разной скоростью. Задача спортсмена ускориться на повороте и плавно выйти на прямую. Сложность задачи возрастает по мере уменьшения радиуса поворота.
Ускорение на прямой со входом в поворот
Тренировочная дистанция — 80-100 м. Ускорения выполняются с разной скоростью. Задача спортсмена — ускориться на прямой и плавно войти в поворот. Сложность задачи возрастает по мере уменьшения радиуса поворота.
Бег по малому кругу на разных скоростях
Тренировочная дистанция — 10-50 м. Ускорения выполняются с разной скоростью. Задача спортсмена отработать технику кругового движения под разным углом, начиная с бега по кругу в 50 м, постепенно уменьшая его радиус до 10 м.
Равномерный бег по повороту
Тренировочная дистанция — 60-80 м. Задача спортсмена в спокойном равномерном темпе — отрабатывать бег по повороту, начиная с крайних дорожек, постепенно доходя до 1 и 2 дорожки.
Имитация работы рук на повороте
Это упражнение выполняется стоя на одном месте. Работают только руки. Все движения выполняются плавно и ритмично.
Количество повторений каждого из упражнений зависит от подготовленности спортсмена — оптимально начинать с выполнения трёх повторений, постепенно увеличивая до 8-10.
Выполняя эти простые упражнения, вы сможете усовершенствовать свою технику бегу, что поможет максимально эффективно и экономично с точки зрения энергозатрат преодолевать любые дистанции за меньший отрезок времени.
Фото: «Фармэко — Бегом по Золотому кольцу», Станислав Зайнуллин
© ГБПОУ КК ПАТИС
ГБПОУ КК ПАТИС
Государственное бюджетное профессиональное образовательное учреждение Краснодарского края
Приморско-Ахтарский техникум индустрии и сервиса
Адрес: 353860 г. Приморско-Ахтарск, ул. Тамаровского, 85
Приморско-Ахтарск, ул. Тамаровского, 85
тел: 8 (861-43) 2-35-94, 8 (861-43) 2-18-98
Адрес сайта: http://патис.рф
Социальные сети: VK и OK
Электронная почта: patiskk@mail.ru
Режим работы:
ПН — СБ: с 8.00 до 16.00
Выходные дни: ВС
Учредители
Наименование:
Министерство образования, науки и молодежной политики Краснодарского края
Адрес: 350063 г. Краснодар, ул. Рашпилевская, 23
тел: 8 (861) 298-25-73
Адрес сайта: minobr.krasnodar.ru
Электронная почта: minobrkuban@krasnodar.ru
Режим работы:
ПН.ВТ.СР.ЧТ. – с 09.00 до 18.00
ПТ. – с 09.00 до 17.00
Перерыв на обед: с 13. 00 до 13.50
00 до 13.50
Выходные дни: СБ.ВС.
Наименование:
Департамент имущественных отношений Краснодарского края
Адрес: 350000 г. Краснодар, ул. Гимназическая, 36
Канцелярия: 8 (861) 268-24-08
Факс: 8 (861) 267-11-75
Специалист по работе с обращениями граждан — консультации, запись на прием — телефон 267-11-78
Телефон горячей линии по вопросам земельных отношений: 8 (861) 992-33-35
Адрес сайта: diok.krasnodar.ru
Электронная почта: dio@krasnodar.ru
Режим работы:
ПН.ВТ.СР.ЧТ. – с 09.00 до 18.00
ПТ. – с 09.00 до 17.00
Перерыв на обед ПН.ВТ.СР.ЧТ.: с 13.00 до 13.50
Перерыв на обед ПТ.: с 13.00 до 13.40
Выходные дни: СБ. ВС.
ВС.
Какой темп бега считается правильным, отличие темпа и скорости
Новички обычно сталкиваются с проблемой по поводу определения темпа бега. Грамотно распределить свои силы на всю дистанцию – это половина успеха. Поэтому так важно знать какой темп бега считается правильным, и давать правильную нагрузку на организм.
Какой темп бега считается правильным
Темп бега будет отличаться в зависимости от дистанции и физической подготовки бегуна. Однако, существует ряд критериев, которые определяют правильность скорости для той или иной дистанции:
- Пульс. Это лучший индикатор того, что спортсмен бежит правильно. Во время легкой пробежки пульс не должен превышать 140 ударов в минуту. При темповом кроссе пульс может повышаться до 180 ударов, но при таких значениях продолжать тренировку можно только в случае полной уверенности в здоровье сердца. Если же такой уверенности нет, то пульс во время бега не рекомендуется поднимать выше 150 ударов в минуту.
- Дыхание. Оно должно быть спокойным и равномерным. Если спортсмен начинает чувствовать, что кислорода недостаточно и дыхание сбивается, значит бег происходит на грани возможностей. Так можно бежать тогда, когда делается финишный рывок, или когда дистанция не превышает 3 км и пробежать ее надо на максимуме. Во всех остальных случаях сбой в дыхании – это признак того, что очень скоро наступит усталость, и тогда темп бега придется снижать до минимума.
- Зажатость. Новички быстро устают и начинают сжимать кулаки и поднимать плечи. Если это ваш вариант, значит вы бежите только за счет силы воли. Темп нужно подбирать таким образом, что не было надобности вынужденно контролировать свою зажатость.
- Приседание. Когда темп бега высокий, а до конца дистанции еще далеко, бегуны экономят силы и приседают к земле. Такая техника, наоборот, приводит к тому, что энергии на работу ног затрачивается больше. Кроме того, дополнительной энергии требует и увеличение частоты шагов. Если ноги спортсмена сильные, но выносливости не хватает, такой бег еще может считаться оправданным, во всех других случаях мышцы ног быстро забьются молочной кислотой.
- Раскачивание головой и телом. Это верный признак усталости, и если продолжать бег в заданном темпе, сил до конца дистанции может не хватить. Однако, некоторые спортсмены по какой-то причине выбирают именно такую технику бега. Поэтому этот критерий надо брать в расчет только если вы уверены, что это не ваша техника.


В чем отличие
Техника бега, дыхание, пульс, нагрузка на мышцы и другие показатели отличаются в зависимости от темпа бега.
Бег быстрым темпом
Быстрый бег обладает большим количеством преимуществ. Он позволяет добиться высокой скорости, выносливости, самореализации, координации и психологической выдержки.
Обычно у спортсменов, тренирующих выносливость, частота пульса во время быстрого бега поднимается до 145-165 ударов в минуту. При этой частоте происходит адаптация организма, поскольку аэробный порог превышается. Если тренировки рассчитаны на скорость, пульс может повыситься до 180 ударов в минуту, но, как уже говорилось, с таким пульсом может бегать только абсолютно здоровый спортсмен.
Во время быстрого бега организму нужно больше кислорода, поэтому вдох должен быть глубоким, а при выдохе нужно полностью выталкивать отработанный воздух из легких.
Справка! Рекомендуется использовать «нижнее» дыхание – сначала заполняется нижняя часть легких, затем верхняя. Такая техника дыхания позволит избежать покалывание в боку.
Основным фактором в быстром беге является техника, так как только за счет нее можно улучшить свои скоростные показатели. Во время марафона техника позволяет правильно распределить свои силы, и увеличить КПД, в спринте – добиться лучшего результата.
Техника быстрого бега:
- Положение рук – руки должны быть согнуты в локтях, кисти расслаблены, движение рук должно происходить в такт бега. Руки помогают ногам, и позволяют развивать большую скорость, затрачивая меньше сил.
- Длина шага – это индивидуальной показатель, он зависит от длины конечностей, роста и уровня физической подготовки. Слишком большие шаги снижают скорость, так как в этом случае стопа опережает центр тяжести. Когда происходит приземление на стопу, нужно контролировать ее положение – она должна находиться под коленом. Корпус остается прямым, взгляд – перед собой. Ошибкой является опускание головы, так как в этом случае весь шейный отдел напрягается.
 Слишком большие шаги снижают скорость, так как в этом случае стопа опережает центр тяжести. Когда происходит приземление на стопу, нужно контролировать ее положение – она должна находиться под коленом. Корпус остается прямым, взгляд – перед собой. Ошибкой является опускание головы, так как в этом случае весь шейный отдел напрягается.
Слишком большие шаги снижают скорость, так как в этом случае стопа опережает центр тяжести. Когда происходит приземление на стопу, нужно контролировать ее положение – она должна находиться под коленом. Корпус остается прямым, взгляд – перед собой. Ошибкой является опускание головы, так как в этом случае весь шейный отдел напрягается.Средний темп бега
Благодаря сниженному темпу бега нагрузка на суставы снижается, следовательно, риск травм уменьшается. Во время среднего бега задействуются все мышцы тела, и сердечная в том числе.
Справка! Нагрузка при среднем темпе должна быть достаточной. Если она слишком низкая, сердечная мышца не будет полноценно качать кровь.
Пульс во время среднего темпа бега составляет 120-140 ударов в минуту. Дыхание должно быть таким, чтобы бегун могу поддерживать разговор и при этом оно не сбивалось. Между вдохом и выдохом нужно делать от двух до четырех шагов.
Бег в медленном темпе
Медики говорят, что медленный темп бега для здоровья гораздо полезнее, чем быстрый. Медленный темп бега подразумевает скорость 7-9 км/час.
Медленный темп бега подразумевает скорость 7-9 км/час.
Дыхание во время легкого бега спокойное, ровное. Движения выполняются без напряжения, легко. Бегун может легко поддерживать разговор и при этом не задыхаться.
Что касается пульса, его диапазон равняется 120-150 ударов в минуту.
Легкий бег тренирует мышцы, но больше всего он полезен для сердца.
Как повысить свои аэробные возможности
Аэробная выносливость – это то, что тренируют тысячи спортсменов, занимающихся разными видами спорта. Задача – максимально отодвинуть аэробный порог, то есть ту точку, после которой в мышцах начинается образование молочной кислоты.
Вот варианты увеличения аэробных возможностей:
Темп и скорость бега
Темп бега – это значение, противоположное средней скорости. Оно измеряется в км/мин и показывает за какое время бегун пробежал ту или иную дистанцию. То есть, если контролировать данный параметр можно рассчитать сколько времени понадобится бегуну на преодоление дистанции.
Скорость бега зависит от веса и роста человека, а также от выносливости организма и поставленной цели.
Обычный человек, который тренируется для здоровья, может пробежать стометровку за 13-15 секунд. Средняя беговая скорость равна у мужчин 15-20 км/ч, у женщин – 12-15 км/ч.
Высокая скорость для здоровья не нужна, она необходима только для установки рекордов.
Резкое увеличение темпа бега
Отрыв или резкое увеличение темпа бега необходимо перед самым финишем. В это время бегун должен бежать на максимальной скорости и на пределе своих возможностей.
Лучше всего финишное ускорение тренируется короткими скоростными интервалами, которые длятся от 30 сек до 2 минут. Тренировка проводится следующим образом:
- разминка;
- 10 отрезков по 400 метров.
Медленный бег будет занимать от 30 сек до 2-3 минут в зависимости от тренированности бегуна. То есть бегун должен бежать 400 метров с максимальной скоростью, затем он переходит на медленный бег, и опять делает рывок на максимальной скорости.
Такая тренировка считается достаточно трудной для выполнения. Как вариант можно бегать 100, 200, 300 метров и сокращать количество повторов.
Советы от профессионалов
Для профессиональных спортсменов существует специальный калькулятор темпа бега, но для бегунов невысокого класса он бесполезен. Бег – это не математика, отслеживать свои пробежки можно и при помощи спортивных часов.
Для тех, кто хочет увеличить темповый параметр, рекомендуется:
- продумывать программу тренировок, включать в них упражнения на выносливость;
- придумать себе хороший мотивационный фактор;
- не заниматься при физическом и нервном истощении;
- приобрести спортивную экипировку;
- бегать в комфортных погодных условиях;
- развивать мышцы ног;
- увеличивать частоту и длину шага;
- бегать на короткие дистанции;
- соблюдать правильную технику бега;
- помнить, что темп бега измеряется в км, значит вы должны выдерживать длительные дистанции, и улучшать при этом временные показатели.

От бега нужно получать удовольствие. Ставьте перед собой реальные цели, и тогда хорошее настроение и положительный настрой помогу вам пройти путь от дивана до 10-километровой пробежки и даже установить собственный рекорд.
Вконтакте
Google+
Speed Training — 10 преимуществ для здоровья бегунов
Разница между средним бегуном и лучшим бегуном не в километраже. И это не вес, диета, обувь или психологическая устойчивость. Это скорость ног. Но скоростная тренировка — это не просто быстрый бег на 5 км. Повышение темпа также обеспечит целый ряд других преимуществ для фитнеса и здоровья. Как будто более быстрого времени недостаточно, вот 10 подтвержденных экспертами причин, по которым вам следует добавить больше скоростной работы в свою рутину.
1.Более сильный шаг
Скорость вашего бега определяется шагом, а именно тем, как далеко вы переносите каждый шаг и как быстро вы можете его выполнить. Есть две фазы шага: время, которое вы проводите в воздухе, и время, когда вы находитесь на земле.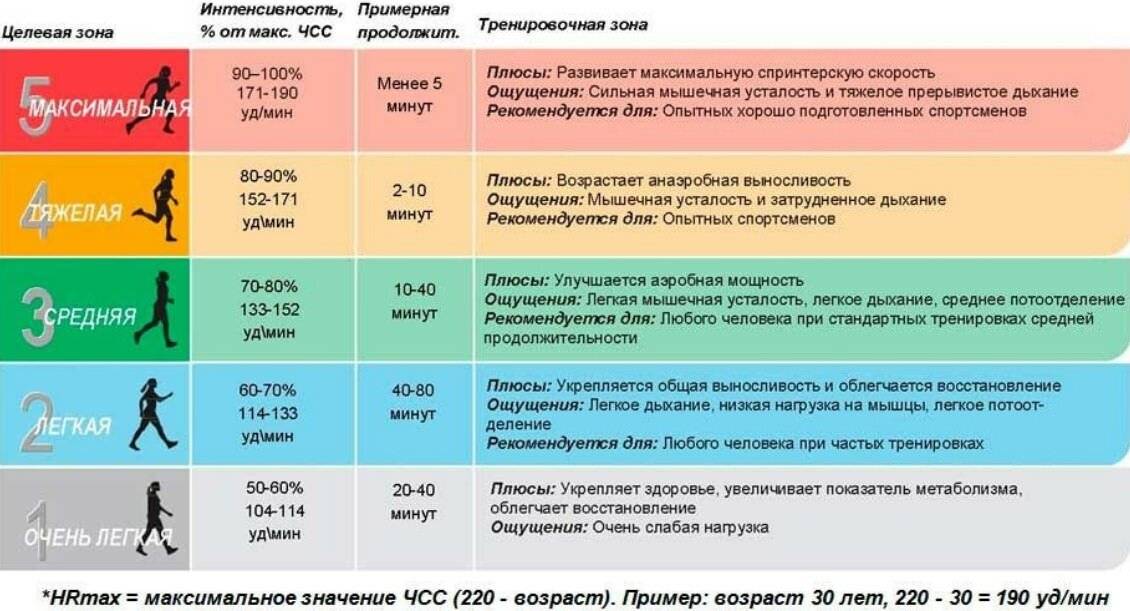 На земле вы прикладываете достаточно силы, чтобы вернуться в воздух. В воздухе вы меняете положение ног для следующего приземления. А теперь самое удивительное: все, от Усэйна Болта до последнего места, занявшего местную 5-километровую дистанцию, могут одинаково быстро менять положение ног в воздухе.Это означает, что разница в скорости ног определяется тем, что происходит на земле. Бегуны, которые быстро прикладывают большее усилие к земле, быстрее возвращаются в воздух и делают более длинный шаг. Конечно, вы не сделаете этого, топая ногами, как сердитый малыш. Вместо этого вы должны тренировать свою нервную систему и мышцы, чтобы увеличивать выработку силы с каждым шагом. Попробуйте такие упражнения, как приседания с прыжком, упражнения с прыжками и прыжки в глубину, чтобы разогреть ноги.
На земле вы прикладываете достаточно силы, чтобы вернуться в воздух. В воздухе вы меняете положение ног для следующего приземления. А теперь самое удивительное: все, от Усэйна Болта до последнего места, занявшего местную 5-километровую дистанцию, могут одинаково быстро менять положение ног в воздухе.Это означает, что разница в скорости ног определяется тем, что происходит на земле. Бегуны, которые быстро прикладывают большее усилие к земле, быстрее возвращаются в воздух и делают более длинный шаг. Конечно, вы не сделаете этого, топая ногами, как сердитый малыш. Вместо этого вы должны тренировать свою нервную систему и мышцы, чтобы увеличивать выработку силы с каждым шагом. Попробуйте такие упражнения, как приседания с прыжком, упражнения с прыжками и прыжки в глубину, чтобы разогреть ноги.
2. Больше мышц
Мы начинаем терять мышечную массу примерно в 25 лет.Хорошая новость заключается в том, что медленно сокращающиеся мышечные волокна (научный термин для «мышечных клеток»), на которые вы больше всего полагаетесь во время бега на длинные дистанции, очень устойчивы к возрастной атрофии. Плохая новость заключается в том, что более быстрые мышечные волокна, определяющие длину шага и необходимые для достижения наилучших результатов в гонках от 5 км до марафона, исчезают со скоростью до 1% в год. Результатом является более медленное время и среднее сокращение длины шага на 40 процентов в течение всей вашей жизни. Но не паникуйте. Сочетая тренировки с отягощениями и скоростную работу, вы можете сократить потерю мышечных волокон вдвое.
Плохая новость заключается в том, что более быстрые мышечные волокна, определяющие длину шага и необходимые для достижения наилучших результатов в гонках от 5 км до марафона, исчезают со скоростью до 1% в год. Результатом является более медленное время и среднее сокращение длины шага на 40 процентов в течение всей вашей жизни. Но не паникуйте. Сочетая тренировки с отягощениями и скоростную работу, вы можете сократить потерю мышечных волокон вдвое.
3. Лучшее сжигание жира
С какой скоростью вы получаете 100 процентов своей энергии из жира? Это вопрос с подвохом, потому что ответ таков: вы сжигаете 100% жира, стоя на месте после спринта, упражнений с отягощениями или других высокоинтенсивных упражнений. Когда вы бегаете, только около 70 процентов вашей энергии поступает из жира. Во время бега на дистанции это около 50 процентов. В темпе мили и быстрее вы используете 100 процентов углеводов для энергии. Но после этих высокоинтенсивных усилий ваше тело экономит углеводы, используя 100-процентный жир для пополнения запасов анаэробной системы первой реакции и для восстановления энергии. Так что выходи и беги!
Так что выходи и беги!
4. Снижение риска травм
Верно. Тренировка на скорость не увеличивает риск получения травмы; это уменьшает его — если вы делаете это правильно. Скоростные тренировки позволяют вашим мышцам выполнять более полный диапазон движений, улучшая гибкость. Он тренирует больше мышц (и больше мышечных волокон внутри мышц), что приводит к лучшему мышечному балансу. И он включает в себя упражнения, которые напрямую укрепляют склонные к травмам мышцы. Возьмем, к примеру, подколенные сухожилия. Ваши подколенные сухожилия выдерживают нагрузки, в 8-10 раз превышающие массу тела, непосредственно перед и после того, как ваша нога коснется земли.Выполняя упражнения, которые укрепляют подколенные сухожилия на этом этапе, вы снижаете шансы получить травму подколенного сухожилия на две трети.
5. Более крепкие кости и другие соединительные ткани
Так же, как мышцы, кости и сухожилия являются живой тканью, поэтому они реагируют на тренировку, становясь сильнее. Исследование, проведенное в Стэнфорде в 2008 году, в котором приняли участие почти тысяча бегунов и не бегунов, пришло к выводу, что бегунам реже требуется замена коленного или тазобедренного сустава. Добавляя скоростную работу и тренировку с отягощениями, вы можете усилить укрепляющий эффект бега для костей, сухожилий, фасций и даже связок.
Исследование, проведенное в Стэнфорде в 2008 году, в котором приняли участие почти тысяча бегунов и не бегунов, пришло к выводу, что бегунам реже требуется замена коленного или тазобедренного сустава. Добавляя скоростную работу и тренировку с отягощениями, вы можете усилить укрепляющий эффект бега для костей, сухожилий, фасций и даже связок.
6. Улучшенная экономия бега
Экономия бега — сложная концепция, но отчасти она измеряет, насколько эффективно вы используете кислород при заданной скорости бега. Используйте свой текущий темп на 5 км в качестве примера. Если вы улучшите экономичность бега, вам потребуется меньше кислорода для бега в таком темпе. С запасным кислородом вы можете либо ускорить темп, либо сохранить текущий темп для более длительной гонки, например, 8 км или 10 км. Высокоинтенсивные тренировки являются ключом к улучшению компонента нервной системы в экономичном беге, и многочисленные исследования показывают улучшение до 6 процентов всего за четыре-шесть недель.
7. Больше анаэробной выносливости
Многие бегуны ошибочно полагают, что бег — за исключением мили на время или финишного толчка в гонке — является исключительно аэробным. Не правда. Первые 30-40 секунд любого бега или гонки в значительной степени подпитываются анаэробной энергией. В начале тренировки есть 30–40-секундная задержка, прежде чем вы сможете обеспечить свои мышцы достаточным количеством кислорода для увеличения выработки аэробной энергии (вся аэробная энергия создается в ваших мышцах). До тех пор ваша анаэробная система заполняет этот пробел.Выполняя скоростную работу, вы уменьшите усталость, связанную с производством анаэробной энергии.
8. Улучшение равновесия и проприоцепции
Без равновесия вы будете падать каждый раз, когда ваша нога коснется земли. А без проприоцепции (осознания положения вашего тела и движения в пространстве) ваши ноги спутывались бы при каждом шаге. К сожалению, бег на длинные дистанции обеспечивает минимальную тренировку этих двух чувств. Исследование, проведенное в 2013 году, показало, что бегуны, пересекающие шатающуюся платформу, внезапно теряли равновесие.Их нервная система и мышцы просто отключаются. Включив в свои скоростные тренировки динамические движения, такие как бег назад, боковые шаги и упражнения на одной ноге, вы улучшите свою способность справляться с любой местностью.
Исследование, проведенное в 2013 году, показало, что бегуны, пересекающие шатающуюся платформу, внезапно теряли равновесие.Их нервная система и мышцы просто отключаются. Включив в свои скоростные тренировки динамические движения, такие как бег назад, боковые шаги и упражнения на одной ноге, вы улучшите свою способность справляться с любой местностью.
9. Улучшенная ловкость
Ловкость предназначена для футболистов, комбайнов НФЛ и детских игр вроде классиков, верно? Неправильный. Ловкость помогает преодолевать крутые повороты на тропе, спрыгивать с бордюра и уворачиваться от собак, которые мчатся на поводке. Ловкость зависит от баланса и проприоцепции, но также требует собственного уникального рецепта нервно-мышечной коммуникации и укрепления.Выполнение быстрой работы ног с помощью упражнений на лестнице и конусе также повысит вашу ловкость после бега.
10. Легкие шаги
Каждый раз, когда ваша нога приземляется, ваши сухожилия и другие соединительные ткани растягиваются, накапливая энергию, вырабатываемую движением и гравитацией. Через долю секунды вы высвобождаете эту энергию — как рогатка, которая катапультирует вас обратно в воздух. Более 50 процентов энергии, используемой во время каждого шага, приходится на упругую отдачу, поэтому укрепление задействованных сухожилий и проводящих путей нервной системы сделает вас сильнее, быстрее и сделает ваш шаг более легким.
Через долю секунды вы высвобождаете эту энергию — как рогатка, которая катапультирует вас обратно в воздух. Более 50 процентов энергии, используемой во время каждого шага, приходится на упругую отдачу, поэтому укрепление задействованных сухожилий и проводящих путей нервной системы сделает вас сильнее, быстрее и сделает ваш шаг более легким.
Вывод: тренировка скорости — это больше, чем просто скорость. И если вы бегун на длинные дистанции, который не тренируется на скорость и силу, вас проигрывает тот, кто тренируется.
Амазонка
Пит Мэджилл — писатель, тренер, бегун и 5-кратный бегун года на турнире Masters Cross Country в США. Адаптировано с разрешения SpeedRunner Пита Мэджилла. Купить сейчас Этот контент создается и поддерживается третьей стороной и импортируется на эту страницу, чтобы помочь пользователям указать свои адреса электронной почты.Вы можете найти дополнительную информацию об этом и подобном контенте на сайте piano. io.
io.
Вот как именно ваш рост влияет на вашу скорость
Несмотря на то, что существует множество способов максимизировать свои беговые результаты, есть и другие факторы, которые вы не можете контролировать, например, ваш рост.
Темп у всех разный, и это во многом может быть связано с длиной ног.
Если ваши ноги короче, вам может показаться, что вам сложнее угнаться за человеком с более длинными конечностями.Но не исключено, что это ощущение просто в твоей голове.
Мы поговорили с тренером Aaptiv Келли Чейз, которая не позволяет своему росту мешать ей достигать таких высоких скоростей, чтобы узнать, как рост влияет на скорость.
Влияет ли рост на скорость бега?
На самом деле нет — поверь самой Чейз. «Лично я невысокого роста. Мой рост 5 футов 2 дюйма, и я никогда не думал, что смогу развивать более высокую скорость при беге, и я считаю, что это настоящий миф, в который верит большинство людей: «Я невысокий, я никогда не смогу бегать так же быстро, как кто-то, кто высокий. ’”
’”
На самом деле это не так уж важно, как думают люди. На самом деле, хотя у высоких людей может быть более длинная длина шага, они также обычно несут больший вес, чем более низкие люди, что может уравнять игровое поле (хотя это, конечно, варьируется от человека к человеку).
Другие факторы, влияющие на скорость бега: гибкость и сила.
Независимо от того, какой у вас рост, бег для увеличения скорости требует и того, и другого. Даже самый высокий человек в мире не смог бы достичь своего максимального темпа, если бы он не занимался силовыми тренировками или не улучшал гибкость.Aaptiv может помочь с нашими тренировками по растяжке!
Так что не позволяйте высоте помешать вам в погоне за этими быстрыми временами. «Я думал так же, но, честно говоря, после непрерывных тренировок с использованием упражнений, которые бросали вызов моему разуму и телу, таких как повторения в гору, спринтерская работа [и] даже просто интервалы прогрессии темпа, мой темп значительно улучшился», — говорит Чейз.
Сосредоточьтесь на последовательных и разнообразных тренировках, чтобы достичь максимальной скорости.
Как улучшить свой темп
Когда дело доходит до быстрого бега, есть много других переменных, которые имеют гораздо большее значение, чем ваш рост.Вот несколько способов улучшить скорость вашего темпа без лишних дюймов.
Поднимитесь на ступеньку выше.
Охватывайте различные типы беговых тренировок, чтобы вы могли воспользоваться преимуществами каждого из них. По словам Чейза, бег в гору особенно полезен для развития скорости.
Дополнительное сопротивление от наклона помогает работать и укреплять силу шага. По мере того, как вы пробиваетесь вверх, ваши ноги привыкают к дополнительному сопротивлению.
Попробуйте пробежаться по ровной местности после изнурительного подъема.Это намного проще, и вы будете двигаться намного быстрее.
Включите беговые тренировки на наклонной поверхности примерно два раза в неделю в свою тренировочную программу. Вы можете делать это на беговой дорожке или найти холмистую местность на улице.
Вы можете делать это на беговой дорожке или найти холмистую местность на улице.
Работайте над своей выносливостью.
Скорость нужна не только для быстрых рывков, таких как спринты. Для бегунов на длинные дистанции более быстрый темп может означать разницу между попаданием в личный рекорд или его отсутствием.
Итак, Чейз рекомендует также сосредоточиться на выносливости. «Вы можете и улучшите свою выносливость и форму шага, что приведет к увеличению темпа», — говорит она.
Неважно, высокий вы или низкий, если у вас нет большой выносливости, вы будете быстро выгорать и замедляться. Улучшив этот аспект, вы получите больше энергии для бега и дольше будете поддерживать скорость.
Добавьте более длительную пробежку на выносливость или пробежку с более длинными (2-3 минуты) интервалами в постоянном или темповом темпе, чтобы повысить выносливость и силу.
Сосредоточьтесь на интервалах увеличения темпа.
Неважно, высокий вы или низкий, вы можете улучшить свои беговые качества — ваш рост не ограничен.
И интервалы прогрессии — ваш лучший друг. Для этого Чейз рекомендует начинать пробежку в удобном темпе.
На такой скорости вы сможете поддерживать разговор и дышать комфортно. Оттуда постепенно увеличивайте темп на 0,2 каждые несколько минут.
Вы можете разогнаться до определенной скорости, например темпа темпа, удерживать ее в течение двух минут, а затем снова снижать скорость.
Помните, что прогресс на самом деле заключается в медленном построении.Если вы напрягаетесь слишком сильно или слишком быстро, то вы увеличиваете риск получения травмы, которая может на время приостановить вашу тренировку или замедлить ее еще больше.
«Ваши мышцы не привыкли к этим более высоким скоростям, [то есть] до тех пор, пока они не будут хорошо подготовлены для них», — говорит Чейз.
Чейз сама испытала на себе воздействие нагрузки на мышцы, когда они не были готовы. «На самом деле у меня был тендинит ахиллова сухожилия из-за отсутствия надлежащей разминки и растяжки, а также из-за того, что я бросала себе вызов слишком быстро, вместо того, чтобы делать это медленно и прогрессировать до более высоких скоростей», — объясняет она.
Так что не спеши и не забывай разогреваться и остывать! Если вы потратите время перед пробежкой, чтобы подготовить свое тело, это поможет вам лучше провести тренировку.
Aaptiv может помочь вам добиться наилучших результатов с нашими фитнес-тренировками на основе звука. Зарегистрируйтесь и сэкономьте сегодня!
Зависит ли выживание от того, насколько быстро вы можете бегать или как быстро вы бегаете? — HUSAK — 2006 — Функциональная экология
Введение
Целями естественного и полового отбора обычно являются фенотипические признаки, представляющие собой интеграцию более низких уровней биологической организации.Следовательно, эволюция морфологических и физиологических признаков обусловлена отбором функциональных признаков более высокого уровня, которые ограничивают то, что животное фактически делает в природе (Huey & Stevenson, 1979; Arnold, 1983; Pough, 1989; Bennett & Huey, 1990). Эта парадигма мышления побудила многих исследователей выдвинуть и эмпирически проверить гипотезы, связывающие индивидуальные различия в приспособленности с индивидуальными различиями в максимальной способности выполнять экологически значимые задачи (т. е. производительность всего животного, Huey & Stevenson 1979; Arnold 1983; Emerson & Arnold 1989). .Соответственно, такие признаки, как максимальная двигательная активность, часто предсказывают выживание, но не для всех видов или демографических групп внутри вида (например, Jayne & Bennett, 1990; Le Galliard, Clobert & Ferrière, 2004; Husak, 2006). Однако отсутствие связи между максимальной производительностью и выживанием не обязательно означает, что производительность не важна для выживания или не является целью отбора.
е. производительность всего животного, Huey & Stevenson 1979; Arnold 1983; Emerson & Arnold 1989). .Соответственно, такие признаки, как максимальная двигательная активность, часто предсказывают выживание, но не для всех видов или демографических групп внутри вида (например, Jayne & Bennett, 1990; Le Galliard, Clobert & Ferrière, 2004; Husak, 2006). Однако отсутствие связи между максимальной производительностью и выживанием не обязательно означает, что производительность не важна для выживания или не является целью отбора.
«Поведение» было предложено многими авторами в качестве фильтра между производительностью и фитнесом (рассмотрено в Garland, Bennett & Daniels, 1990; Garland & Losos, 1994; Irschick & Garland, 2001), однако связи между производительностью, поведением и фитнесом остаются неясными, в основном из-за отсутствия эмпирических данных и неполной теоретической основы для работы (Webb 1986; Bennett 1989; Garland & Losos 1994; Kingsolver et al . 2001 г.; Иршик и др. . 2005). Гипотетические направленные связи предполагают, что максимальная производительность создает верхний предел того, на что способен организм, но при котором он может использовать набор различных поведенческих вариантов (Bennett 1989; Garland 1994; Garland & Losos 1994). Текущие гипотезы также предполагают, что максимальная мощность определяет, что делает организм при выполнении задачи в природе. Например, максимально быстрые особи должны убегать от хищников быстрее, чем максимально медленные особи.
2001 г.; Иршик и др. . 2005). Гипотетические направленные связи предполагают, что максимальная производительность создает верхний предел того, на что способен организм, но при котором он может использовать набор различных поведенческих вариантов (Bennett 1989; Garland 1994; Garland & Losos 1994). Текущие гипотезы также предполагают, что максимальная мощность определяет, что делает организм при выполнении задачи в природе. Например, максимально быстрые особи должны убегать от хищников быстрее, чем максимально медленные особи.
Эти проблемы еще более усложняются, поскольку не существует четкого функционального различия между «поведением» и «производительностью» (см. обзоры Irschick & Garland 2001; Lailvaux & Irschick 2006). Связь между производительностью и «поведением» легче количественно изучить, когда интересующей поведенческой чертой является использование максимальной производительности в природе (т. е. «экологическая производительность»; Irschick 2003). Например, с какой скоростью люди бегают в природе по сравнению с максимальной скоростью, с которой они могут бегать в лаборатории? Учитывая этот пример, «производительность» может быть определена в традиционном смысле как максимальная способность организма выполнять экологическую задачу с использованием динамического движения (т.г. кусать, бегать, летать или плавать), а «экологическую эффективность» можно определить как то, как организм использует максимальную производительность в природе при выполнении экологической задачи (Irschick 2003). «Поведение» — гораздо более широкий термин, описывающий действия организма, но не дающий количественную оценку того, насколько хорошо он выполняет данную задачу (обзор см. в Lailvaux & Irschick, 2006). Примером, позволяющим различать эти три, является реакция бегства животного. Он имеет способность убегать с некоторой скоростью, ограниченной его морфологией и физиологией («производительность»), но после оценки ситуации убегает со скоростью ниже своих максимальных возможностей («экологическая производительность»).
Например, с какой скоростью люди бегают в природе по сравнению с максимальной скоростью, с которой они могут бегать в лаборатории? Учитывая этот пример, «производительность» может быть определена в традиционном смысле как максимальная способность организма выполнять экологическую задачу с использованием динамического движения (т.г. кусать, бегать, летать или плавать), а «экологическую эффективность» можно определить как то, как организм использует максимальную производительность в природе при выполнении экологической задачи (Irschick 2003). «Поведение» — гораздо более широкий термин, описывающий действия организма, но не дающий количественную оценку того, насколько хорошо он выполняет данную задачу (обзор см. в Lailvaux & Irschick, 2006). Примером, позволяющим различать эти три, является реакция бегства животного. Он имеет способность убегать с некоторой скоростью, ограниченной его морфологией и физиологией («производительность»), но после оценки ситуации убегает со скоростью ниже своих максимальных возможностей («экологическая производительность»). Акт «ответа на хищника» и есть «поведение». Из-за широты и неточности термина «поведение», используемого другими в дискуссиях о морфологической эволюции, я впредь использую «экологическое поведение» вместо «поведения» и призываю будущих исследователей различать эти два понятия в своих собственных исследованиях с признанием что оба, вероятно, важны для эволюции морфологических признаков.
Акт «ответа на хищника» и есть «поведение». Из-за широты и неточности термина «поведение», используемого другими в дискуссиях о морфологической эволюции, я впредь использую «экологическое поведение» вместо «поведения» и призываю будущих исследователей различать эти два понятия в своих собственных исследованиях с признанием что оба, вероятно, важны для эволюции морфологических признаков.
Передвижение ящерицы послужило полезной модельной системой в эволюционной биологии, особенно в исследованиях отбора по максимальным признакам производительности (обзор в Garland & Losos 1994; Aerts et al .2000 г.; Иршик и Гарланд, 2001). Максимальная скорость бега предсказывает выживание вылупившихся ящериц у некоторых видов (Warner & Andrews 2002; Miles 2004; Husak 2006), но не у других (см. Bennett & Huey 1990). Однако предсказательная сила максимальной скорости бега на выживание взрослых ящериц, демографической группы, прошедшей по крайней мере один раунд отбора, остается малоизученной. Максимальная скорость спринта не предсказывала выживание взрослых кольчатых ящериц ( Crotaphytus Collaris ), но предсказывала выживание вылупившихся детенышей (Husak 2006), предполагая, что существуют онтогенетические изменения в роли естественного отбора в эволюции локомоторных характеристик (см. также Jayne и Беннетт, 1990).Кроме того, то, как и когда ящерицы используют максимальную скорость бега, зависит от экологического контекста: особи некоторых возрастов и полов быстро бегают, спасаясь от хищников или преследуя соперников, но не во время кормления (Irschick & Losos 1998; Irschick 2000; Irschick et al . 2000; Маттингли и Джейн, 2004; Хусак и Фокс, 2006). Эти наблюдения повышают вероятность того, что экологические показатели, а не максимальные показатели, могут быть целью отбора для некоторых видов или демографических групп внутри вида.Несмотря на способность экологических показателей предсказывать выживание, насколько мне известно, ни одно исследование не проверяло, что лучше предсказывает выживание: максимальная производительность или экологические показатели.
Максимальная скорость спринта не предсказывала выживание взрослых кольчатых ящериц ( Crotaphytus Collaris ), но предсказывала выживание вылупившихся детенышей (Husak 2006), предполагая, что существуют онтогенетические изменения в роли естественного отбора в эволюции локомоторных характеристик (см. также Jayne и Беннетт, 1990).Кроме того, то, как и когда ящерицы используют максимальную скорость бега, зависит от экологического контекста: особи некоторых возрастов и полов быстро бегают, спасаясь от хищников или преследуя соперников, но не во время кормления (Irschick & Losos 1998; Irschick 2000; Irschick et al . 2000; Маттингли и Джейн, 2004; Хусак и Фокс, 2006). Эти наблюдения повышают вероятность того, что экологические показатели, а не максимальные показатели, могут быть целью отбора для некоторых видов или демографических групп внутри вида.Несмотря на способность экологических показателей предсказывать выживание, насколько мне известно, ни одно исследование не проверяло, что лучше предсказывает выживание: максимальная производительность или экологические показатели.
Я проверил силу отбора на максимальную продуктивность и экологическую результативность в двух контекстах у взрослых кольчатых ящериц, широко распространенного вида на юго-западе США. Ошейниковые ящерицы используют сидение и ожидание кормления, чтобы по возможности питаться доступными членистоногими (Blair & Blair 1941; Cooper et al .2001), и на них охотятся самые разные хищники (Husak et al . 2006). Следовательно, побег хищника и поимка добычи могут быть контекстами, в которых максимальная скорость спринта подвергается отбору. И наоборот, поскольку кольчатые ящерицы в среднем не используют максимальную способность спринта при поиске пищи или бегстве от хищников (Husak & Fox 2006), скорость, с которой ящерицы выполняют эти задачи, может подвергаться отбору. С другой стороны, абсолютные скорости, используемые в природе, не могут быть объектом отбора; вместо этого доля максимальной мощности, используемой в природе во время определенных задач, может быть целью отбора. Поэтому я проверил, является ли максимальная скорость спринта, скорость при поиске пищи или скорость при бегстве от хищников предсказателями выживания, и зависит ли выживание от возраста или пола. Затем я проверил, зависит ли выживание от пропорции максимальной мощности, используемой в природе при поиске пищи или побеге от хищников.
Поэтому я проверил, является ли максимальная скорость спринта, скорость при поиске пищи или скорость при бегстве от хищников предсказателями выживания, и зависит ли выживание от возраста или пола. Затем я проверил, зависит ли выживание от пропорции максимальной мощности, используемой в природе при поиске пищи или побеге от хищников.
Материалы и методы
выживание
Исследование проводилось на плотине Сунер-Лейк в округе Пауни, штат Оклахома, где субстрат состоял из покрытых бетоном валунов.В апреле-мае 2003 г. я поймал взрослых (≥ 2 лет) и годовалых (≈ 1 года) самцов и самок ящериц, надев на них петли и навсегда пометив их, обрезав пальцы и нанеся уникальный узор из цветных пятен на лапах. задние конечности для визуальной идентификации на расстоянии (модификация Baird, Acree & Sloan 1996). Чтобы избежать влияния обрезки пальцев на двигательную активность, я никогда не обрезал самый длинный палец ни у одной ящерицы. Скорость спринта у людей определяли перед подрезанием пальцев в начале сезона и снова в конце сезона после подрезания пальцев, и не было существенной разницы в максимальной скорости спринта (J.Гусак (неопубликованные данные). Особи, пойманные мной в предыдущем году до появления птенцов, считались возрастом ≥ 2 лет. Я исследовал выживаемость 60 взрослых и годовалых ящериц (14 годовалых самок, 15 годовалых самцов, 18 самок ≥ 2 лет и 13 самцов ≥ 2 лет). Ящерицы, проанализированные в этом исследовании, представляют собой другую выборку той же популяции, что и исследованная в Husak (2006).
Скорость спринта у людей определяли перед подрезанием пальцев в начале сезона и снова в конце сезона после подрезания пальцев, и не было существенной разницы в максимальной скорости спринта (J.Гусак (неопубликованные данные). Особи, пойманные мной в предыдущем году до появления птенцов, считались возрастом ≥ 2 лет. Я исследовал выживаемость 60 взрослых и годовалых ящериц (14 годовалых самок, 15 годовалых самцов, 18 самок ≥ 2 лет и 13 самцов ≥ 2 лет). Ящерицы, проанализированные в этом исследовании, представляют собой другую выборку той же популяции, что и исследованная в Husak (2006).
Я определял выживаемость в полевых условиях до начала следующего сезона активности, проводя ежедневные учеты исследуемого участка и прилегающих к нему участков подходящей среды обитания.Время между выпуском и оценкой выживания охватывало два экологически важных периода, в течение которых ящерицы могли погибнуть: (1) в период активности (примерно конец марта – начало августа), когда хищничество, голод или травмы, связанные с внутри- или межвидовыми взаимодействиями, могут вызвали гибель и (2) во время зимовки (приблизительно с середины августа до конца марта), когда ящерицы находятся в состоянии покоя и когда смертность могла быть вызвана замерзанием, хищничеством, инфекцией или недостатком запасов энергии. Моя цель, однако, заключалась в том, чтобы определить, какие признаки лучше всего предсказывают выживание в течение всего года, а не предсказывать выживание в течение одного или другого из двух экологических периодов. Предыдущая работа с популяцией показала, что ни размер тела, ни состояние тела (или другие измеренные морфологические переменные) не предсказывают выживаемость годовалых и взрослых кольчатых ящериц (Husak 2006), что позволило мне сосредоточить свой анализ в этом исследовании на переменных производительности, которые потенциально важны.
Моя цель, однако, заключалась в том, чтобы определить, какие признаки лучше всего предсказывают выживание в течение всего года, а не предсказывать выживание в течение одного или другого из двух экологических периодов. Предыдущая работа с популяцией показала, что ни размер тела, ни состояние тела (или другие измеренные морфологические переменные) не предсказывают выживаемость годовалых и взрослых кольчатых ящериц (Husak 2006), что позволило мне сосредоточить свой анализ в этом исследовании на переменных производительности, которые потенциально важны.
максимальная скорость бега
90 002 ящерицы были доставлены в Государственный университет Оклахомы и задержаны на 1 день, чтобы обеспечить прохождение содержимого кишечника.Беременных самок не использовали. По утрам на второй и третий дни их содержания в неволе я помещал ящериц по отдельности в сетчатые мешки в освещенный инкубатор при температуре 37 °C. Трижды в день с интервалом между испытаниями не менее 2 ч каждую ящерицу вынимали из инкубатора и сразу же отгоняли по 3-метровой беговой дорожке в черный мешок. Беговая дорожка была покрыта наждачной бумагой с высоким коэффициентом трения. Я записал прогоны на пленку SONY Hi8 (после Braña 2003), а позже просмотрел ленты и подсчитал количество кадров, необходимых для прохождения 1-метровых интервалов (стабильность записи скорости кадров для используемой камеры и лент была проверена путем записи секундомер в отдельном эксперименте Дж.Гусак (неопубликованные данные). Полученные скорости были сравнимы со скоростями, полученными Peterson & Husak (2006) при использовании беговой дорожки с инфракрасными лучами. Я классифицировал качество каждого прогона как «хорошее» или «плохое» (van Berkum & Tsuji, 1987), а данные о «плохих» прогонах (паузы, реверсы) отбрасывали. Анализировался только один самый быстрый шпагат на 1 м для каждого забега. После пробежки я сразу же проверил температуру тела ящерицы с помощью быстрого клоакального термометра. Все ящерицы были выпущены в точном месте их отлова после того, как все испытания были завершены в конце третьего дня их содержания в неволе.
Беговая дорожка была покрыта наждачной бумагой с высоким коэффициентом трения. Я записал прогоны на пленку SONY Hi8 (после Braña 2003), а позже просмотрел ленты и подсчитал количество кадров, необходимых для прохождения 1-метровых интервалов (стабильность записи скорости кадров для используемой камеры и лент была проверена путем записи секундомер в отдельном эксперименте Дж.Гусак (неопубликованные данные). Полученные скорости были сравнимы со скоростями, полученными Peterson & Husak (2006) при использовании беговой дорожки с инфракрасными лучами. Я классифицировал качество каждого прогона как «хорошее» или «плохое» (van Berkum & Tsuji, 1987), а данные о «плохих» прогонах (паузы, реверсы) отбрасывали. Анализировался только один самый быстрый шпагат на 1 м для каждого забега. После пробежки я сразу же проверил температуру тела ящерицы с помощью быстрого клоакального термометра. Все ящерицы были выпущены в точном месте их отлова после того, как все испытания были завершены в конце третьего дня их содержания в неволе. Лучшие и вторые лучшие результаты каждого человека, независимо от дня, в который они произошли, были сильно коррелированы, что указывает на высокую повторяемость ( r = 0,90, P < 0,001).
Лучшие и вторые лучшие результаты каждого человека, независимо от дня, в который они произошли, были сильно коррелированы, что указывает на высокую повторяемость ( r = 0,90, P < 0,001).полевых скоростей
Для определения производительности в полевых условиях я использовал методы Irschick & Losos (1998) и Husak & Fox (2006). Я измерил скорость бега для всех половозрастных классов в разгар сезона размножения в двух случаях: (1) бегство от хищника и (2) поиск пищи.В этих испытаниях не использовались беременные самки. Для всех контекстов использовалась одна и та же обобщенная процедура получения скорости спринта. Я использовал видеомагнитофон, чтобы записать движения ящериц, а затем просмотрел запись в поле, чтобы использовать ориентиры для измерения расстояния, пройденного для каждого движения. Я подсчитал количество кадров (записано 30 кадров с 90 175 – 1 90 176) на единицу расстояния, измеренного в поле, чтобы получить скорость спринта. Дистанции, пробежанные у разных людей, различались, поэтому для анализа использовались только пробежки ≥ 3 м, чтобы их можно было сравнить с лабораторными измерениями максимальной скорости.Скорости рассчитывались для приблизительных последовательных интервалов в 1 м (среднее ± SE = 1,2 ± 0,3 м), чтобы облегчить сравнение этих скоростей со скоростями, измеренными в лаборатории. Для анализа мы использовали самый быстрый приближенный 1-метровый интервал. Если во время испытания происходило несколько движений, я использовал самую быструю скорость. Все испытания проводились при температуре субстрата 30–40 °C, что является оптимальным температурным диапазоном для кольчатых ящериц в центральной части Оклахомы (Uzee 1990).
Дистанции, пробежанные у разных людей, различались, поэтому для анализа использовались только пробежки ≥ 3 м, чтобы их можно было сравнить с лабораторными измерениями максимальной скорости.Скорости рассчитывались для приблизительных последовательных интервалов в 1 м (среднее ± SE = 1,2 ± 0,3 м), чтобы облегчить сравнение этих скоростей со скоростями, измеренными в лаборатории. Для анализа мы использовали самый быстрый приближенный 1-метровый интервал. Если во время испытания происходило несколько движений, я использовал самую быструю скорость. Все испытания проводились при температуре субстрата 30–40 °C, что является оптимальным температурным диапазоном для кольчатых ящериц в центральной части Оклахомы (Uzee 1990).
В ходе испытаний по побегу хищника человек (в каждом случае один и тот же человек, одетый в одинаковую одежду) шел к ящерице с постоянной скоростью примерно 40 м/мин −1 (согласно Cooper 1997).К ящерице приближались параллельно направлению плотины, чтобы ящерицы не бежали вверх или вниз по плотине, наклон которой может повлиять на поведенческие стратегии борьбы с хищниками (Jayne & Ellis, 1998) и спринтерские характеристики (Irschick & Jayne, 1999).
Чтобы вызвать движения при кормлении, я вызвал попытку поиска пищи, привязав муху (без крючка) к концу 4-метрового шеста и поместив ее на расстоянии 3 м от ящерицы, слегка покачивая ее в том месте, на котором она находилась. размещен (т.г. Иршик 2000; Хусак и Фокс, 2006 г.). Я использовал это расстояние, чтобы его можно было сравнить с расстоянием, используемым в лаборатории для измерения максимальной производительности. Движения ящериц по направлению к мухе записывались на видео, и скорости рассчитывались описанным выше способом.
статистический анализ
Сначала я использовал двусторонний t -критерий для изучения различий в чертах между выжившими и не выжившими. Я использовал логистическую регрессию (т.г. Janzen & Stern 1998) для проверки как направленного (линейного), так и стабилизирующего или разрушительного (квадратичного) отбора (Lande & Arnold 1983). Все независимые переменные были логарифмически 10 преобразованы для анализа. Зависимой переменной была непреобразованная вероятность выживания (т.е. 0 или 1). Модель логистической регрессии линейного отбора включала максимальную скорость спринта, скорость спринта при поиске пищи, скорость спринта при бегстве от хищника, возраст (взрослый = 1, годовалый = 0) и пол (самец = 1, самка = 0).Квадратичная модель логистической регрессии отбора включала квадратичные значения для каждого из трех измерений скорости спринта. Там, где были получены значимые модели логистической регрессии, использовались методы Janzen & Stern (1998) для преобразования стандартизированных (т.е. умноженных на стандартное отклонение признака перед отбором, σ z ) коэффициентов логистической регрессии в коэффициенты отбора с использованием средний градиент оцениваемой поверхности отбора (т.е. вероятности выживания).В результате коэффициенты отбора (β avggrad ) сравнимы с коэффициентами, полученными при множественной линейной регрессии (Janzen & Stern 1998).
Зависимой переменной была непреобразованная вероятность выживания (т.е. 0 или 1). Модель логистической регрессии линейного отбора включала максимальную скорость спринта, скорость спринта при поиске пищи, скорость спринта при бегстве от хищника, возраст (взрослый = 1, годовалый = 0) и пол (самец = 1, самка = 0).Квадратичная модель логистической регрессии отбора включала квадратичные значения для каждого из трех измерений скорости спринта. Там, где были получены значимые модели логистической регрессии, использовались методы Janzen & Stern (1998) для преобразования стандартизированных (т.е. умноженных на стандартное отклонение признака перед отбором, σ z ) коэффициентов логистической регрессии в коэффициенты отбора с использованием средний градиент оцениваемой поверхности отбора (т.е. вероятности выживания).В результате коэффициенты отбора (β avggrad ) сравнимы с коэффициентами, полученными при множественной линейной регрессии (Janzen & Stern 1998).
Отбор может основываться на абсолютных скоростях, с которыми ящерицы бегают в природе, или на доле максимальной используемой мощности. Следовательно, для каждой особи я разделил каждую скорость поля на соответствующую максимальную скорость, чтобы получить пропорции максимальной скорости спринта, используемой при поиске пищи и побеге.Затем я выполнил второй набор логистических регрессионных анализов (как линейных, так и квадратичных, как указано выше), чтобы определить, является ли пропорция максимальной способности при поиске пищи или побеге предсказанным выживанием. Все анализы проводились в SAS по сравнению с 9·1 (SAS Institute Inc., Кэри, Северная Каролина, США).
Следовательно, для каждой особи я разделил каждую скорость поля на соответствующую максимальную скорость, чтобы получить пропорции максимальной скорости спринта, используемой при поиске пищи и побеге.Затем я выполнил второй набор логистических регрессионных анализов (как линейных, так и квадратичных, как указано выше), чтобы определить, является ли пропорция максимальной способности при поиске пищи или побеге предсказанным выживанием. Все анализы проводились в SAS по сравнению с 9·1 (SAS Institute Inc., Кэри, Северная Каролина, США).
Результаты
Из 60 ящериц, помеченных в 2003 г. и включенных в это исследование, 33 (55%) дожили до 2004 г. Изучив каждую демографическую группу, я обнаружил, что восемь (44%) взрослых самок, шесть годовалых самок (67%), 13 взрослых самцов ( выжили 65%) и шесть годовалых самцов (46%).Выжившие и не выжившие существенно не различались по максимальной скорости спринта ( t = 0,51, df = 58, P = 0,61, таблица 1) или скорости во время поиска пищи ( t = 0,50 , df = 58, P = 0·62, таблица 1). Однако выжившие бежали быстрее, спасаясь от хищников, чем невыжившие ( t = 4·46, d.f. = 43·7, P < 0·001, Таблица 1). Анализ распределения полевых скоростей показал, что скорости, используемые во время двух действий, очень мало перекрываются (рис.1а), особенно для особей, доживших до следующего года (рис. 1б).
Однако выжившие бежали быстрее, спасаясь от хищников, чем невыжившие ( t = 4·46, d.f. = 43·7, P < 0·001, Таблица 1). Анализ распределения полевых скоростей показал, что скорости, используемые во время двух действий, очень мало перекрываются (рис.1а), особенно для особей, доживших до следующего года (рис. 1б).
| Скорость бега (мс −1 ) | Выжившие (33) | Невыжившие (27) |
|---|---|---|
| Максимальная | 3·38 ± 0·07 | 3·44 ± 0·09 |
| Собирательство | 1·11 ± 0·05 | 1,08 ± 0,06 |
| Побег | 2·47 ± 0·04 | 2·14 ± 0·06 |
Частотные распределения скоростей, используемых в природе до отбора при поиске пищи (сплошные столбцы) и побеге от моделируемого хищника (незаштрихованные столбцы) для (а) всех особей и (б) тех особей, которые дожили до следующего года.
Модель логистической регрессии линейного отбора с максимальной и полевой скоростями в качестве независимых переменных была значимой (χ 2 = 21·31, df = 5, P < 0·001), при этом только скорость убегающего спринта достоверно предсказывала выживаемость (Таблица 2 ). Модель логистической регрессии квадратичного отбора также была значимой (χ 2 = 23,39, df = 8, P = 0,003), но ни один из коэффициентов частичной регрессии не был значимым ( P > 0,2 для все), предполагая, что значимая модель была обусловлена наличием скорости ускользающего спринта.В совокупности эти результаты указывают на то, что направленный отбор основан только на скорости спринтерского бегства, а не на стабилизирующем или разрушительном отборе. Результаты качественно одинаковы, если используются остаточные значения (см. также Husak 2006) или если годовики и взрослые ящерицы анализируются отдельно, а самцы и самки анализируются отдельно.
| Коэффициент выбора | P -значение | |
|---|---|---|
| Возраст | −0·084 | 0·30 |
| Секс | 0·090 | 0·32 |
| Максимальная скорость бега | 0·079 | 0·45 |
| Скорость спринта при поиске пищи | 0·063 | 0·53 |
| Скорость побега | 0·383 | < 0·001 |
Модель логистической регрессии линейного отбора с долей максимальной мощности, используемой при поиске пищи и побеге в качестве независимых переменных, была значимой (χ 2 = 11·93, d.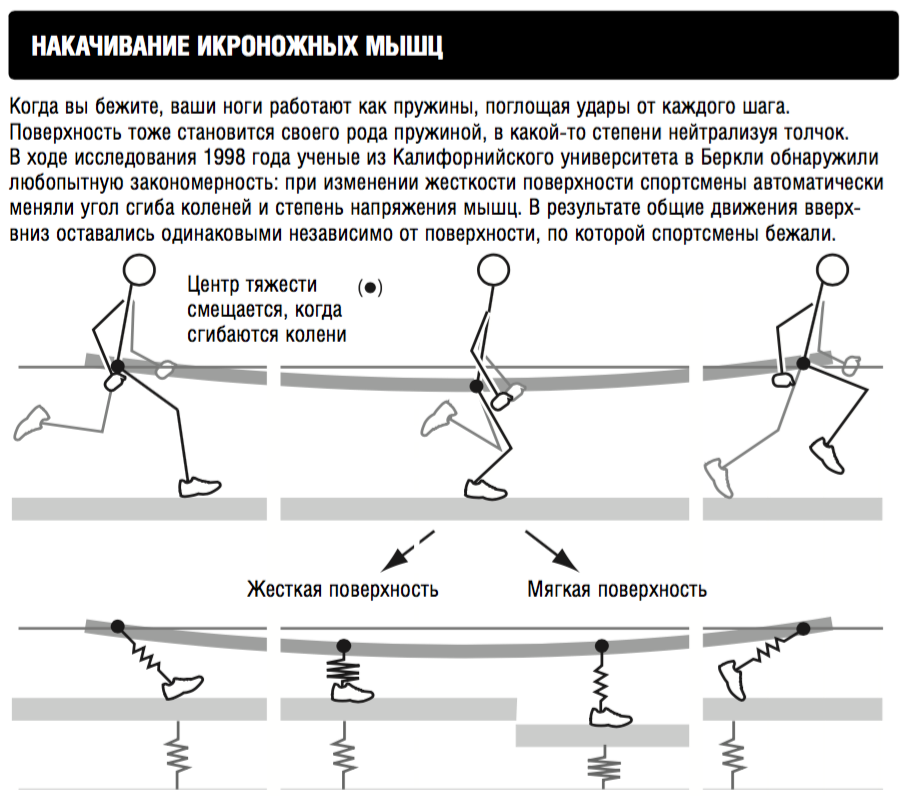 ф. = 2, P = 0·003), причем доля максимальной емкости, используемой при побеге, является значимой (β avggrad = 0,316, P = 0,002), а доля максимальной емкости, используемой при поиске пищи, является незначительно значимой (β avggrad = -0·173, P = 0·052). Модель логистической регрессии квадратичного отбора также была значимой (χ 2 = 12·12, df = 4, P = 0,016), но ни один из коэффициентов частичной регрессии не был значимым ( P > 0,5 для все), предполагая, что значимая модель была обусловлена наличием переменных, значимых в линейной модели.В совокупности эти результаты указывают на направленный отбор доли максимальной мощности, используемой при выполнении экологических задач, но в разных направлениях, в зависимости от контекста. Результаты качественно совпадают, если анализировать отдельно годовиков и взрослых ящериц, а также отдельно анализировать самцов и самок.
ф. = 2, P = 0·003), причем доля максимальной емкости, используемой при побеге, является значимой (β avggrad = 0,316, P = 0,002), а доля максимальной емкости, используемой при поиске пищи, является незначительно значимой (β avggrad = -0·173, P = 0·052). Модель логистической регрессии квадратичного отбора также была значимой (χ 2 = 12·12, df = 4, P = 0,016), но ни один из коэффициентов частичной регрессии не был значимым ( P > 0,5 для все), предполагая, что значимая модель была обусловлена наличием переменных, значимых в линейной модели.В совокупности эти результаты указывают на направленный отбор доли максимальной мощности, используемой при выполнении экологических задач, но в разных направлениях, в зависимости от контекста. Результаты качественно совпадают, если анализировать отдельно годовиков и взрослых ящериц, а также отдельно анализировать самцов и самок.
Обсуждение
Результаты этого исследования показали, что отбор может воздействовать на экологические показатели, подчеркивая, что исследования, направленные на лучшее выяснение целей отбора, должны учитывать как сами максимальные признаки продуктивности всего животного, так и экологические показатели. Следовательно, исследования, которые не обнаруживают связи между производительностью и выживанием, должны учитывать актуальность этой характеристики производительности в природе, чтобы окончательно определить роль этой характеристики производительности в эволюции лежащей в ее основе морфологии. Если отбор не опирается на максимальную производительность, возможно, он действует на экологическую производительность, которая по определению определяется и ограничивается максимальной производительностью.
Следовательно, исследования, которые не обнаруживают связи между производительностью и выживанием, должны учитывать актуальность этой характеристики производительности в природе, чтобы окончательно определить роль этой характеристики производительности в эволюции лежащей в ее основе морфологии. Если отбор не опирается на максимальную производительность, возможно, он действует на экологическую производительность, которая по определению определяется и ограничивается максимальной производительностью.
Скорость, с которой годовалые и взрослые кольчатые ящерицы убегают от хищников, по-видимому, важнее для выживания, чем максимальная скорость, с которой они могут бегать.Это отличается от вылупившихся кольчатых ящериц, у которых выживание определяется максимальной скоростью бега (Husak 2006). Судьба вылупившихся детенышей, вероятно, зависит от их максимальных спринтерских способностей, а не от их поведенческих решений из-за их ограниченных спринтерских способностей по сравнению со старшими (и, соответственно, более крупными) особями (Carrier 1996; Husak 2006; Herrel & Gibb 2006). Годовалые и взрослые особи, которые могут бегать намного быстрее, чем детеныши, обладают большей гибкостью в стратегии, которую они используют для спасения от хищников.Действительно, кольчатые ящерицы старше 2 лет используют меньше своих максимальных спринтерских способностей при бегстве от хищников, чем годовалые (Husak & Fox 2006), а это означает, что с возрастом кольчатые ящерицы могут развивать скорость, необходимую для спасения от хищников (т.е. или «адекватная» скорость убегания; см. Husak & Fox 2006), используя меньше своей максимальной мощности. Это онтогенетическое изменение экологических характеристик поддерживает идею о том, что максимальная производительность обеспечивает «пространство производительности», в котором ограничиваются поведенческие решения животного (Bennett 1989).По мере того, как ящерицы стареют и увеличиваются в размерах, это пространство, вероятно, увеличивается, что позволяет выразить больше возможностей для экологических характеристик. Эти гипотезы предполагают более сложный сценарий предполагаемого просачивающегося эффекта отбора на экологические показатели, чем если бы вместо этого он действовал только на основе максимальной производительности.
Годовалые и взрослые особи, которые могут бегать намного быстрее, чем детеныши, обладают большей гибкостью в стратегии, которую они используют для спасения от хищников.Действительно, кольчатые ящерицы старше 2 лет используют меньше своих максимальных спринтерских способностей при бегстве от хищников, чем годовалые (Husak & Fox 2006), а это означает, что с возрастом кольчатые ящерицы могут развивать скорость, необходимую для спасения от хищников (т.е. или «адекватная» скорость убегания; см. Husak & Fox 2006), используя меньше своей максимальной мощности. Это онтогенетическое изменение экологических характеристик поддерживает идею о том, что максимальная производительность обеспечивает «пространство производительности», в котором ограничиваются поведенческие решения животного (Bennett 1989).По мере того, как ящерицы стареют и увеличиваются в размерах, это пространство, вероятно, увеличивается, что позволяет выразить больше возможностей для экологических характеристик. Эти гипотезы предполагают более сложный сценарий предполагаемого просачивающегося эффекта отбора на экологические показатели, чем если бы вместо этого он действовал только на основе максимальной производительности. Если скорость побега — это то, на что действует отбор, то ожидается эволюционное изменение скелетно-мышечной системы двигательного аппарата, а также сенсорных систем, связанных с обнаружением хищника, и нервных путей для обработки такой информации и принятия «соответствующих» поведенческих решений (Лаудер). и Лием, 1989; Лаудер и Рейли, 1996).Крайне необходимы исследования, которые стремятся непосредственно связать все эти переменные.
Если скорость побега — это то, на что действует отбор, то ожидается эволюционное изменение скелетно-мышечной системы двигательного аппарата, а также сенсорных систем, связанных с обнаружением хищника, и нервных путей для обработки такой информации и принятия «соответствующих» поведенческих решений (Лаудер). и Лием, 1989; Лаудер и Рейли, 1996).Крайне необходимы исследования, которые стремятся непосредственно связать все эти переменные.
Несмотря на то, что более быстро убегающие воротниковые ящерицы выживали лучше, чем те, которые убегали медленнее, более быстрые убегающие не обязательно являются максимально быстрыми особями. Не существует положительной связи между скоростью побега и максимальной скоростью спринта (Husak & Fox 2006), предполагая, что максимальная способность ограничивает , но не обязательно определяет то, что люди в конечном итоге делают, реагируя на хищника.Это также подтверждается тем фактом, что более быстрые беглецы не были более быстрыми собирателями ( r = 0,24, P = 0,07), хотя связь имеет положительную тенденцию. Интересно, что существует значимая отрицательная корреляция между скоростью убегания хищника и расстоянием, на которое воротничковые ящерицы позволяют приблизиться симулированному хищнику (т. е. «расстояние сближения», рассчитанное на основе наших испытаний по спасению хищников, согласно Cooper 1997; r = -0·63, P < 0·001), так что медленно убегающие особи не позволяют хищникам приближаться так же близко, как более быстрые убегающие.Это подразумевает разные стратегии побега от хищников среди кольчатых ящериц, и почти бимодальное распределение скоростей побега до отбора (рис. 1а) подтверждает это наблюдение. Однако не было обнаружено подрывного отбора по скорости побега или пропорции максимальной мощности, используемой при побеге. Нет существенной связи между скоростью убегания и расстоянием, которое кольчатые ящерицы пробегают после приближения симулированного хищника (т. е. «расстояние полета», рассчитанное на основе наших испытаний по спасению хищников; согласно Cooper 1997; r = 0·10, P = 0,64), ни между максимальной скоростью спринта и любой переменной антихищника ( P = 0,23 для расстояния приближения и P = 0,72 для расстояния полета).
Интересно, что существует значимая отрицательная корреляция между скоростью убегания хищника и расстоянием, на которое воротничковые ящерицы позволяют приблизиться симулированному хищнику (т. е. «расстояние сближения», рассчитанное на основе наших испытаний по спасению хищников, согласно Cooper 1997; r = -0·63, P < 0·001), так что медленно убегающие особи не позволяют хищникам приближаться так же близко, как более быстрые убегающие.Это подразумевает разные стратегии побега от хищников среди кольчатых ящериц, и почти бимодальное распределение скоростей побега до отбора (рис. 1а) подтверждает это наблюдение. Однако не было обнаружено подрывного отбора по скорости побега или пропорции максимальной мощности, используемой при побеге. Нет существенной связи между скоростью убегания и расстоянием, которое кольчатые ящерицы пробегают после приближения симулированного хищника (т. е. «расстояние полета», рассчитанное на основе наших испытаний по спасению хищников; согласно Cooper 1997; r = 0·10, P = 0,64), ни между максимальной скоростью спринта и любой переменной антихищника ( P = 0,23 для расстояния приближения и P = 0,72 для расстояния полета). Однако, несмотря на корреляцию между скоростью убегания и расстоянием сближения, когда расстояние сближения включено в логистическую регрессию с проанализированными выше переменными производительности, расстояние сближения не является значимым предиктором выживания (β = 0·22, P = 0· 38). Эти результаты подчеркивают наше непонимание того, как максимальная продуктивность влияет на экологическую результативность, и последствия поведенческих решений, основанных на максимальных возможностях, на фитнесе.
Однако, несмотря на корреляцию между скоростью убегания и расстоянием сближения, когда расстояние сближения включено в логистическую регрессию с проанализированными выше переменными производительности, расстояние сближения не является значимым предиктором выживания (β = 0·22, P = 0· 38). Эти результаты подчеркивают наше непонимание того, как максимальная продуктивность влияет на экологическую результативность, и последствия поведенческих решений, основанных на максимальных возможностях, на фитнесе.
Обнаружение того, что люди, использующие более высокую долю своей максимальной скорости при побеге, выживали лучше, чем те, кто использовал меньше, было несколько неожиданным, поскольку использование почти максимальной скорости может быть стрессовым или энергетически вредным.Поскольку доля максимальной мощности, используемой в природе, рассчитывается как отношение, положительная связь между выживанием и долей максимальной мощности, используемой при побеге, может быть получена тремя способами. Во-первых, вероятность выживания может увеличиваться по мере увеличения скорости убегания и максимальной вместимости, но при этом скорость ускользания увеличивается быстрее, чем максимальная вместимость. Во-вторых, вероятность выживания может увеличиваться, поскольку скорость убегания остается постоянной, но максимальная вместимость уменьшается. В-третьих, вероятность выживания может увеличиваться по мере увеличения скорости побега, но максимальная вместимость не имеет отношения к выживанию.Селекционные анализы максимальной и полевой скоростей не подтверждают первые две возможности, так как не было связи между максимальной способностью и выживаемостью, и была положительная связь между выживаемостью и скоростью побега. Третья возможность согласуется с анализом отбора и поддерживает гипотезу о том, что выживание зависит от достижения некоторой скорости, достаточной для побега от хищников, независимо от максимальной способности. Третий сценарий также подразумевает, что положительная связь между выживанием и долей максимальной мощности, используемой при побеге, является артефактом скорости побега, являющейся числителем при расчете пропорции.
Во-первых, вероятность выживания может увеличиваться по мере увеличения скорости убегания и максимальной вместимости, но при этом скорость ускользания увеличивается быстрее, чем максимальная вместимость. Во-вторых, вероятность выживания может увеличиваться, поскольку скорость убегания остается постоянной, но максимальная вместимость уменьшается. В-третьих, вероятность выживания может увеличиваться по мере увеличения скорости побега, но максимальная вместимость не имеет отношения к выживанию.Селекционные анализы максимальной и полевой скоростей не подтверждают первые две возможности, так как не было связи между максимальной способностью и выживаемостью, и была положительная связь между выживаемостью и скоростью побега. Третья возможность согласуется с анализом отбора и поддерживает гипотезу о том, что выживание зависит от достижения некоторой скорости, достаточной для побега от хищников, независимо от максимальной способности. Третий сценарий также подразумевает, что положительная связь между выживанием и долей максимальной мощности, используемой при побеге, является артефактом скорости побега, являющейся числителем при расчете пропорции. Однако мои результаты трудно интерпретировать в сравнительном смысле, потому что немногие исследователи провели количественную оценку отбора по максимальной производительности, и, насколько мне известно, ни один из них не проводил количественной оценки отбора по экологическим характеристикам производительности, которые могут быть ограничены максимальной производительностью (Irschick & Garland 2001; Kingsolver et al. al . 2001). Будущие эмпирические и теоретические исследования должны попытаться выйти за рамки концептуально простой (но сложной с точки зрения логистики) парадигмы морфология-производительность-фитнес и связанных с ней индивидуальных связей.Если не максимальная производительность, а экологическая производительность предсказывает выживание, то механизмы, которые непосредственно регулируют использование максимальной производительности, должны быть исследованы как звенья на более ранних этапах цепи. В будущей работе следует также рассмотреть, как внешние факторы одновременно влияют на морфологию, производительность и экологическую эффективность, и какие последствия эти эффекты имеют на каждом уровне биологической организации и, в конечном итоге, приспособленности.
Однако мои результаты трудно интерпретировать в сравнительном смысле, потому что немногие исследователи провели количественную оценку отбора по максимальной производительности, и, насколько мне известно, ни один из них не проводил количественной оценки отбора по экологическим характеристикам производительности, которые могут быть ограничены максимальной производительностью (Irschick & Garland 2001; Kingsolver et al. al . 2001). Будущие эмпирические и теоретические исследования должны попытаться выйти за рамки концептуально простой (но сложной с точки зрения логистики) парадигмы морфология-производительность-фитнес и связанных с ней индивидуальных связей.Если не максимальная производительность, а экологическая производительность предсказывает выживание, то механизмы, которые непосредственно регулируют использование максимальной производительности, должны быть исследованы как звенья на более ранних этапах цепи. В будущей работе следует также рассмотреть, как внешние факторы одновременно влияют на морфологию, производительность и экологическую эффективность, и какие последствия эти эффекты имеют на каждом уровне биологической организации и, в конечном итоге, приспособленности.
Благодарности
Я хотел бы поблагодарить Оклахома Газ энд Электрик за доступ к месту исследования, П.Виддер, М. Роуз и Н. Расмуссен за помощь в полевых работах, а также С. Фокс, Д. Иршик и К. Лаппин за комментарии к предыдущим версиям рукописи. Эта работа была проведена при финансовой поддержке Национального научного фонда (грант на улучшение диссертации IOB-0407943) и грантов Sigma Xi в поддержку исследований. Все работы проводились в соответствии с разрешением Департамента охраны дикой природы Оклахомы и были одобрены Комитетом по уходу и использованию животных Университета штата Оклахома.
- Артс, П.,
Ван Дамм, Р.,
Ванхойдонк, Б.,
Зааф, А. и
Херрел, А. (2000) Передвижение ящериц: как морфология встречается с экологией. Нидерландский журнал зоологии 50, 261–277.
- Арнольд, С.А. (1983) Морфология, работоспособность и приспособленность. Американский зоолог 23, 347–361.
- Бэрд, Т.А., Акри, Массачусетс и Слоан, К.Л. (1996) Возрастные и гендерные различия в социальном поведении и успехе спаривания свободноживущих ошейниковых ящериц, Crotaphytus collis . Копейя 1996, 336–347.
- Беннет А.Ф. (1989) Комплексные исследования двигательной активности.
Сложные функции организма: интеграция и эволюция позвоночных (ред.
Д.Б. Пробуждение и
Г. Рот), стр. 191–202. John Wiley and Sons, Чичестер, Великобритания.
- Беннетт, А.Ф. и Хьюи, Р. Б. (1990) Изучение эволюции физиологических показателей. Оксфордские обзоры эволюционной биологии 7, 251–284.
- Ван Беркум, Ф.Х. & Цудзи, Дж.С. (1987) Межсемейные различия в скорости бега вылупившихся Sceloporus occidentalis . Журнал зоологии 212, 511– 519.
- Блэр, В.Ф. & Блэр, А. П. (1941) Пищевые привычки ящерицы с воротником на северо-востоке Оклахомы. Американский натуралист Мидленда 26, 230–232.
- Брана, Ф. (2003) Морфологические корреляты скорости взрыва и паттернов движения поля: поведенческая адаптация передвижения у настенных ящериц ( Podarcis muralis ). Биологический журнал Линнеевского общества 80, 135–146.
- Кэрриер, Д. (1996) Онтогенетические ограничения двигательной активности. Физиологическая зоология 69, 467–488.
- Купер, У.Э. Jr (1997) Факторы угрозы, влияющие на антихищное поведение широкоголового сцинка ( Eumeces laticeps ): повторное приближение, изменение пути хищника и зрительный контакт. Копейя 1997, 613–619.
- Купер, У.Э. младший,
Витт, Л. Дж.,
Колдуэлл, Дж. П. и
Фокс, С.Ф. (2001) Режимы кормодобывания некоторых американских ящериц: взаимосвязь между переменными измерения и дискретность режимов. Герпетология 57, 65–76.
- Эмерсон, С.Б. & Арнольд, С.Дж. (1989) Внутри- и межвидовые отношения между морфологией, производительностью и приспособленностью. Сложные функции организма: интеграция и эволюция позвоночных (ред. Д.Б. Пробуждение и Г. Рот), стр. 295–314. John Wiley and Sons, Чичестер, Великобритания.
- Гарланд, Т. младший (1994) Количественная генетика локомоторного поведения и физиологии подвязочной змеи.
Количественные генетические исследования поведенческой эволюции (изд. CRB Boake), стр. 251–277. University of Chicago Press, Чикаго, Иллинойс.
- Гарленд, Т.младший и Лосос, Дж. Б. (1994) Экологическая морфология двигательной активности у чешуйчатых рептилий. Экологическая морфология: интегративная биология организмов (ред. ПК. Уэйнрайт и С.М. Reilly), стр. 240–302. University of Chicago Press, Чикаго, Иллинойс.
- Гарланд, Т. младший, Беннетт, А.Ф. и Дэниелс, CB (1990) Наследуемость двигательной активности и ее корреляты в естественной популяции. Опыт 46, 530–533.
- Херрел, А. и Гибб, AC (2006) Онтогенез производительности у позвоночных. Физиолого-биохимическая зоология 79, 1–6.
- Хьюи, Р. Б. и
Стивенсон, Р. Д. (1979) Объединение тепловой физиологии и экологии эктотермов: обсуждение подходов. Американский зоолог 19, 357–366.
- Хусак, Дж. Ф. (2006) Помогает ли скорость выжить? Тест с ошейниковыми ящерицами разного возраста. Функциональная экология 20, 174–179.
- Хусак, Дж. Ф. и Фокс, С.Ф. (2006) Полевое использование максимальной скорости бега ящерицами с воротником ( Crotaphytus collis ): компенсация и половой отбор. Эволюция 60, 1888–1895 гг.
- Хусак, Дж. Ф.,
Македония, Дж. М.,
Фокс, С.Ф. &
Соседа, Р. К. (2006) Стоимость заметной окраски самцов ящериц с воротником ( Crotaphytus collis ): экспериментальный тест с использованием моделей ящериц, покрытых глиной. Этология 112, 572–580.
- Иршик, Д.J. (2000) Влияние поведения и онтогенеза на двигательную активность западно-индийской ящерицы Anolis lineatopus . Функциональная экология 14, 438–444.
- Иршик, Д.Дж. (2003) Изучение производительности в природе: последствия для вариаций приспособленности в популяциях. Интегративная и сравнительная биология 43, 396–407.
- Иршик, Д.Дж. &
Гарланд, Т. младший (2001) Объединение функции и экологии в исследованиях адаптации: исследования двигательной способности как модельной системы. Ежегодный обзор экологии и систематики 32, 367–396.
- Иршик, Д.Дж. & Джейн, Б.C. (1999) Полевое исследование влияния наклона на побег двуногой ящерицы, Callisaurus draconoides . Физиолого-биохимическая зоология 72, 44– 56.
- Иршик, Д.Дж. & Лосос, Дж. Б. (1998) Сравнительный анализ экологического значения двигательной активности карибских ящериц Anolis . Эволюция 52, 219–226.
- Иршик, Д.Дж.,
Макрини, Т.Е.,
Коруба, С. и
Форман, Дж. (2000) Онтогенетические различия в морфологии, использовании среды обитания, поведении и способности к спринту у двух видов ящериц Вест-Индии Anolis . Герпетологический журнал 34, 444–451.
- Иршик, Д.Дж., Херрел, А., Ванхойдонк, Б., Хьюг, К. и Ван Дамм, Р. (2005) Локомоторная компенсация создает несоответствие между лабораторными и полевыми оценками скорости убегания у ящериц: предостерегающая история для исследований производительности и приспособленности. Эволюция 59, 1579–1587.
- Янзен, Ф.Дж. и Стерн, Х.С. (1998) Логистическая регрессия для эмпирических исследований многомерного отбора. Эволюция 52, 1564–1571.
- Джейн, Британская Колумбия &
Беннетт, А.Ф. (1990) Отбор по двигательным возможностям в естественной популяции подвязочных змей. Эволюция 44, 1204–1229.
- Джейн, Британская Колумбия & Эллис, Р.V. (1998) Как наклоны влияют на поведение ящерицы, обитающей в дюнах, при побеге, Uma scoparia . Поведение животных 55, 1115–1130.
- Кингсолвер, Дж. Г., Хекстра, Х.Е., Хекстра, Дж. М., Берриган, Д., Виньери, С.Н., Хилл, CE, Хоанг, А., Гиберт, П. и Берли, П. (2001) Сила фенотипического отбора в природных популяциях. Американский натуралист 157, 245–261.
- Лейлво, С.П. и
Иршик, Д.Дж. (2006) Функциональный взгляд на половой отбор: идеи и перспективы на будущее. Поведение животных 72, 263–273.
- Ланде, Р. и Арнольд, С.Дж. (1983) Измерение отбора по коррелированным признакам. Эволюция 37, 1210–1226.
- Лаудер, Г.В. & Лием, К.Ф. (1989) Роль исторических факторов в эволюции сложных функций организма. Сложные функции организма: интеграция и эволюция позвоночных (ред. Д.Б. Будить & Г. Рот), стр. 63–78. John Wiley and Sons, Чичестер, Великобритания.
- Лаудер, Г.В. и
Рейли, С.М. (1996) Механистическая основа поведенческой эволюции: сравнительный анализ скелетно-мышечной функции. В: Филогении и сравнительный метод в поведении животных.
(изд. Э. Мартин), стр. 105–137. Издательство Оксфордского университета, Оксфорд.
- Ле Гальяр, Ж.-Ф., Клоберт, Дж. и Ферьер, Р. (2004) Физическая работоспособность и дарвиновская приспособленность у ящериц. Природа 432, 502–505.
- Маттингли, У.Б. & Джейн, Британская Колумбия (2004) Использование ресурсов в древесных местообитаниях: структура влияет на передвижение четырех экоморфов ящериц Anolis . Экология 85, 1111–1124.
- Майлз, Д.Б. (2004) Гонка идет к быстрому: фитнес последствия различий в спринтерских характеристиках у молодых ящериц. Исследования эволюционной экологии 6, 63–75.
- Петерсон, К. С. &
Хусак, Дж. Ф. (2006) Двигательная активность и половой отбор: индивидуальные различия в скорости бега ящериц с воротником ( Crotaphytus collis ). Копейя 2006, 216–224.
- Пуф, Ф.Х. (1989) Производительность организма и дарвиновская приспособленность: подходы и интерпретации. Физиологическая зоология 62, 199–236.
- Uzee, EM (1990) Влияние температурных ограничений на повседневную активность Crotaphytus collis. Неопубликованная магистерская диссертация, Университет штата Оклахома, Стилуотер, Оклахома, США.
- Уорнер, Д.А. &
Эндрюс, Р. М. (2002) Лабораторные и полевые эксперименты определяют источники изменчивости фенотипов и выживаемости вылупившихся ящериц. Биологический журнал Линнеевского общества 76, 105–124.
- Уэбб, П.В. (1986) Передвижение и отношения хищник-жертва. Отношения хищник-жертва: взгляды и подходы из изучения низших позвоночных (ред. М.Э. Федер и Г.В. Lauder), стр. 24–41. University of Chicago Press, Чикаго, Иллинойс.
Каталожные номера
 Нидерландский журнал зоологии 50, 261–277.
Нидерландский журнал зоологии 50, 261–277. Рот), стр. 191–202. John Wiley and Sons, Чичестер, Великобритания.
Рот), стр. 191–202. John Wiley and Sons, Чичестер, Великобритания. (2003) Морфологические корреляты скорости взрыва и паттернов движения поля: поведенческая адаптация передвижения у настенных ящериц ( Podarcis muralis ). Биологический журнал Линнеевского общества 80, 135–146.
(2003) Морфологические корреляты скорости взрыва и паттернов движения поля: поведенческая адаптация передвижения у настенных ящериц ( Podarcis muralis ). Биологический журнал Линнеевского общества 80, 135–146. Дж.,
Колдуэлл, Дж. П. и
Фокс, С.Ф. (2001) Режимы кормодобывания некоторых американских ящериц: взаимосвязь между переменными измерения и дискретность режимов. Герпетология 57, 65–76.
Дж.,
Колдуэлл, Дж. П. и
Фокс, С.Ф. (2001) Режимы кормодобывания некоторых американских ящериц: взаимосвязь между переменными измерения и дискретность режимов. Герпетология 57, 65–76.
 Б. и
Стивенсон, Р. Д. (1979) Объединение тепловой физиологии и экологии эктотермов: обсуждение подходов. Американский зоолог 19, 357–366.
Б. и
Стивенсон, Р. Д. (1979) Объединение тепловой физиологии и экологии эктотермов: обсуждение подходов. Американский зоолог 19, 357–366. К. (2006) Стоимость заметной окраски самцов ящериц с воротником ( Crotaphytus collis ): экспериментальный тест с использованием моделей ящериц, покрытых глиной. Этология 112, 572–580.
К. (2006) Стоимость заметной окраски самцов ящериц с воротником ( Crotaphytus collis ): экспериментальный тест с использованием моделей ящериц, покрытых глиной. Этология 112, 572–580. Ежегодный обзор экологии и систематики 32, 367–396.
Ежегодный обзор экологии и систематики 32, 367–396. Герпетологический журнал 34, 444–451.
Герпетологический журнал 34, 444–451. Эволюция 44, 1204–1229.
Эволюция 44, 1204–1229.

 С. &
Хусак, Дж. Ф. (2006) Двигательная активность и половой отбор: индивидуальные различия в скорости бега ящериц с воротником ( Crotaphytus collis ). Копейя 2006, 216–224.
С. &
Хусак, Дж. Ф. (2006) Двигательная активность и половой отбор: индивидуальные различия в скорости бега ящериц с воротником ( Crotaphytus collis ). Копейя 2006, 216–224. М. (2002) Лабораторные и полевые эксперименты определяют источники изменчивости фенотипов и выживаемости вылупившихся ящериц. Биологический журнал Линнеевского общества 76, 105–124.
М. (2002) Лабораторные и полевые эксперименты определяют источники изменчивости фенотипов и выживаемости вылупившихся ящериц. Биологический журнал Линнеевского общества 76, 105–124.границ | Люди оптимизируют время контакта с землей и жесткость ног, чтобы минимизировать метаболические затраты при беге
Введение
Самооптимизация — это подсознательная тонкая настройка механики бега для минимизации метаболических затрат (Cavanagh and Williams, 1982; Williams and Cavanagh, 1987; Moore et al., 2012, 2016), и считается, что он играет ключевую роль в развитии экономичной беговой походки. Hogberg (1952) представил первый пример систематического управления длиной шага для изучения самооптимизации у тренированного бегуна и сообщения о результирующей метаболической реакции. Кавана и Уильямс (1982) применили эту первоначальную работу к большей группе ( n = 10) тренированных бегунов. В обоих исследованиях наблюдалась криволинейная U-образная зависимость, подчеркивающая, что тренированные бегуны могли самостоятельно выбирать длину шага, которая была равна или близка к их оптимальной длине шага, рассчитанной математически.Другие исследователи воспроизвели эти результаты (Morgan et al., 1994; Hunter and Smith, 2007; de Ruiter et al., 2013; Connick and Li, 2014; van Oeveren et al., 2017) и расширили базу знаний, продемонстрировав, что необученные бегуны дальше от своего математического оптимума, чем тренированные бегуны (de Ruiter et al., 2013), и субмаксимальная скорость бега не влияет на оптимальную длину шага (van Oeveren et al., 2017). Кроме того, даже в состоянии усталости у тренированных бегунов длина шага близка к оптимальной, вычисленной математически (Hunter and Smith, 2007).
Hogberg (1952) представил первый пример систематического управления длиной шага для изучения самооптимизации у тренированного бегуна и сообщения о результирующей метаболической реакции. Кавана и Уильямс (1982) применили эту первоначальную работу к большей группе ( n = 10) тренированных бегунов. В обоих исследованиях наблюдалась криволинейная U-образная зависимость, подчеркивающая, что тренированные бегуны могли самостоятельно выбирать длину шага, которая была равна или близка к их оптимальной длине шага, рассчитанной математически.Другие исследователи воспроизвели эти результаты (Morgan et al., 1994; Hunter and Smith, 2007; de Ruiter et al., 2013; Connick and Li, 2014; van Oeveren et al., 2017) и расширили базу знаний, продемонстрировав, что необученные бегуны дальше от своего математического оптимума, чем тренированные бегуны (de Ruiter et al., 2013), и субмаксимальная скорость бега не влияет на оптимальную длину шага (van Oeveren et al., 2017). Кроме того, даже в состоянии усталости у тренированных бегунов длина шага близка к оптимальной, вычисленной математически (Hunter and Smith, 2007). Следовательно, стало общепризнанным, что длина и частота шагов сами оптимизируются тренированными бегунами. Тем не менее, ограниченное внимание уделялось оценке оптимизации того, как создается частота шагов, в частности учету времени контакта с землей, которое может вызывать различные специфические реакции спортсмена при манипуляциях.
Следовательно, стало общепризнанным, что длина и частота шагов сами оптимизируются тренированными бегунами. Тем не менее, ограниченное внимание уделялось оценке оптимизации того, как создается частота шагов, в частности учету времени контакта с землей, которое может вызывать различные специфические реакции спортсмена при манипуляциях.
Связь между временем контакта с землей и метаболическими затратами была неоднозначной (Moore, 2016). В ранних работах предполагалось, что время контакта с землей обратно пропорционально энергетическим затратам на ходьбу и бег (Kram and Taylor, 1990; Hoyt et al., 1994; Kipp et al., 2018), что означает, что увеличение времени контакта уменьшит энергию, необходимую для перемещения на единицу расстояния. Такую связь наблюдали Ди Микеле и Мерни (2013) и Уильямс и Кавана (1986). Однако с тех пор время, проведенное в контакте с землей, было определено как метаболически затратная фаза цикла ходьбы (Arellano and Kram, 2014), что побудило многих выступать за то, чтобы более короткое время контакта с землей способствовало снижению метаболических затрат (Nummela et al. др., 2007; Сантос-Консехеро и др., 2014 г.; Фолланд и др., 2017). Хотя исследования подтвердили связь с этим эффектом (Nummela et al., 2007; Santos-Concejero et al., 2014, 2017), убедительных причинных доказательств не поступало. Недавно Lussiana et al. (2019) заметили, что короткое и длительное время контакта с землей может быть экономически выгодным в зависимости от типа бегуна. У бегунов, которые проводят относительно большую часть цикла ходьбы в контакте с землей (высокий коэффициент нагрузки), метаболические затраты были такими же, как и у тех, кто проводит относительно небольшую часть цикла ходьбы в контакте с землей (низкий фактор нагрузки) (Lussiana et al. др., 2019). Эти результаты подтверждают теорию самооптимизации, поскольку бегуны, по-видимому, подсознательно адаптировались как механически, так и физиологически. Соответственно, проблемы, связанные с текущими исследованиями, основанными на перекрестных сравнениях, изучающих время контакта и метаболические затраты, означают, что рекомендации для конкретных спортсменов по экономичному бегу и этой конкретной характеристике походки остаются недостижимыми (Moore, 2016).
др., 2007; Сантос-Консехеро и др., 2014 г.; Фолланд и др., 2017). Хотя исследования подтвердили связь с этим эффектом (Nummela et al., 2007; Santos-Concejero et al., 2014, 2017), убедительных причинных доказательств не поступало. Недавно Lussiana et al. (2019) заметили, что короткое и длительное время контакта с землей может быть экономически выгодным в зависимости от типа бегуна. У бегунов, которые проводят относительно большую часть цикла ходьбы в контакте с землей (высокий коэффициент нагрузки), метаболические затраты были такими же, как и у тех, кто проводит относительно небольшую часть цикла ходьбы в контакте с землей (низкий фактор нагрузки) (Lussiana et al. др., 2019). Эти результаты подтверждают теорию самооптимизации, поскольку бегуны, по-видимому, подсознательно адаптировались как механически, так и физиологически. Соответственно, проблемы, связанные с текущими исследованиями, основанными на перекрестных сравнениях, изучающих время контакта и метаболические затраты, означают, что рекомендации для конкретных спортсменов по экономичному бегу и этой конкретной характеристике походки остаются недостижимыми (Moore, 2016).
Морин и др. (2007) были единственными исследователями, которые использовали план с участием одного участника для изучения времени контакта с землей.В частности, они смогли манипулировать временем контакта с землей и продемонстрировали, что изменения времени, проведенного в контакте с землей, объясняют большую долю дисперсии изменений жесткости ног, чем изменения частоты шагов ( r 2 = 0,90 и 0,47). , соответственно). Хотя в исследовании не измерялись метаболические затраты при беге, было обнаружено, что большая жесткость ног связана с более низкими метаболическими затратами (Dalleau et al., 1998) и считается экономичной стратегией бега (Moore, 2016).Следовательно, можно утверждать, что создание большей жесткости ног при одновременном сохранении частоты шагов, чему способствует более короткое время контакта с землей, снизит метаболические затраты при беге. Расчет жесткости ног основан на концепции, согласно которой бег человека можно объяснить с помощью модели массы пружины (Бликхан, 1989). Пружина представляет собой ногу, которая сжимается телом во время первой половины контакта с землей, а затем отскакивает вверх во время второй половины контакта с землей (Morin et al., 2005). Более жесткая нога потенциально будет накапливать и высвобождать энергию более эффективно, чем менее жесткая нога, и впоследствии это может снизить метаболические затраты при беге. Помимо исследования Morin et al. (2007), времени контакта с землей и жесткости ног уделялось ограниченное внимание в исследованиях с участием участников, изучающих экономичный бег.
Пружина представляет собой ногу, которая сжимается телом во время первой половины контакта с землей, а затем отскакивает вверх во время второй половины контакта с землей (Morin et al., 2005). Более жесткая нога потенциально будет накапливать и высвобождать энергию более эффективно, чем менее жесткая нога, и впоследствии это может снизить метаболические затраты при беге. Помимо исследования Morin et al. (2007), времени контакта с землей и жесткости ног уделялось ограниченное внимание в исследованиях с участием участников, изучающих экономичный бег.
Оценка времени контакта бегуна с землей может быть выполнена с помощью относительно простого оборудования, такого как видеокамера или приложение для телефона (Balsalobre-Fernández et al., 2017), благодаря которому биомеханический анализ стал более доступным для тренеров и практиков. Однако для определения оптимального времени контакта с землей с учетом метаболических затрат в настоящее время требуется дорогостоящее оборудование для измерения состава вдыхаемого и выдыхаемого воздуха (например, система газового анализа) и технические знания. Суррогатные меры были эффективно использованы для оценки длины и частоты шагов (de Ruiter et al., 2013), но по-прежнему требуют дополнительного оборудования. Вполне возможно, что рейтинги воспринимаемой нагрузки (RPE) могли бы стать суррогатной, доступной и простой в использовании мерой для изучения воспринимаемой потребности в манипулировании временем контакта с землей.Несколько исследований показали, что метаболические затраты на бег линейно связаны с воспринимаемой нагрузкой (см. обзор Chen et al., 2002), поэтому вполне вероятно, что бег будет эффективным. Кроме того, из-за взаимосвязи между RPE и метаболическими затратами, RPE часто используется тренерами и практиками для мониторинга тренировочных реакций (McLaren et al., 2018), но неизвестно, является ли это подходящей суррогатной мерой для использования в технике. целенаправленное обучение.
Основная цель этого исследования заключалась в изучении влияния манипулирования временем контакта с землей на метаболические затраты при беге, в дополнение к определению влияния измененной жесткости ног на метаболические затраты при беге. Основываясь на теории самооптимизации, было высказано предположение, что тренированные бегуны будут самостоятельно выбирать время контакта с землей и жесткость ног, близкие к их математически определенному метаболически оптимальному времени контакта с землей (в пределах 5%). Математический оптимум представляет собой идентифицируемый минимум в криволинейной зависимости между затратами на метаболизм и характеристиками походки. Второй целью было оценить связь между метаболическими затратами и воспринимаемой нагрузкой в различных условиях времени контакта с землей, и мы предположили, что будет наблюдаться положительная линейная зависимость между метаболической стоимостью и воспринимаемой нагрузкой.
Основываясь на теории самооптимизации, было высказано предположение, что тренированные бегуны будут самостоятельно выбирать время контакта с землей и жесткость ног, близкие к их математически определенному метаболически оптимальному времени контакта с землей (в пределах 5%). Математический оптимум представляет собой идентифицируемый минимум в криволинейной зависимости между затратами на метаболизм и характеристиками походки. Второй целью было оценить связь между метаболическими затратами и воспринимаемой нагрузкой в различных условиях времени контакта с землей, и мы предположили, что будет наблюдаться положительная линейная зависимость между метаболической стоимостью и воспринимаемой нагрузкой.
Методы
Участники
Десять тренированных бегунов на выносливость университетского уровня (девять мужчин и одна женщина) дали информированное письменное согласие на участие в исследовании (возраст: 19,8 ± 2,6 года, рост: 1,79 ± 0,12 м, масса: 65,1 ± 6,6 кг). Каждый участник проходил как минимум две структурированные тренировки в неделю, входил в состав первой команды по легкой атлетике и мог бегать до 17 минут на 5 км или до 35 минут (мужчины) и до 45 минут на 10 км (женщины). Кроме того, все участники были знакомы с беговой дорожкой и не имели травм в течение предыдущих 6 месяцев.Этическое одобрение было получено от Комитета по этике Университета.
Кроме того, все участники были знакомы с беговой дорожкой и не имели травм в течение предыдущих 6 месяцев.Этическое одобрение было получено от Комитета по этике Университета.
Процедура
Все рабочие условия были выполнены во время одного визита в лабораторию. Перед началом разминки измеряли массу и рост. Все участники выполняли самостоятельно выбранную разминку в течение от 5 до 10 минут, в течение которых они были ознакомлены с сигналами, которые должны были быть предоставлены. Затем участников попросили самостоятельно выбрать скорость бега, которую, по их мнению, они смогут комфортно поддерживать в течение 30 минут.Затем участники выполнили 5 х 6-минутных пробежек на беговой дорожке с выбранной ими скоростью бега (12,7 ± 1,6 км·ч -1 ) с 3-минутными периодами отдыха между последовательными подходами. Самостоятельно выбранная скорость бега считалась субмаксимальной на основании данных о привычном состоянии, обеспечивающем коэффициент дыхательного обмена <1,0 в течение последних 2 мин бега. Во время первого забега участники выполняли свою привычную технику бега, что позволяло определить выбранную ими самостоятельно частоту шагов и время контакта с землей через приложение Runmatic .Затем участники выполнили четыре отдельных прогона в стандартном порядке, при этом была предоставлена специфическая словесная подсказка для выявления одного из четырех условий: медленное время контакта, очень медленное время контакта, быстрое время контакта и очень быстрое время контакта. Во время каждого условия метроном использовался для поддержания частоты шагов участников на уровне, соответствующем их самостоятельно выбранной частоте. Конкретные словесные инструкции заключались в том, чтобы «установить контакт с землей в такт ударам метронома и отреагировать на предоставленный сигнал.Вербальный сигнал подавался каждые 30 с (Moore et al., 2019) и заключался в следующем: условие (1) увеличить время контакта больше, чем обычно; условие (2) максимально увеличить время контакта; условие (3) уменьшить время контакта больше, чем обычно и; условие (4) максимально уменьшить время контакта.
Во время первого забега участники выполняли свою привычную технику бега, что позволяло определить выбранную ими самостоятельно частоту шагов и время контакта с землей через приложение Runmatic .Затем участники выполнили четыре отдельных прогона в стандартном порядке, при этом была предоставлена специфическая словесная подсказка для выявления одного из четырех условий: медленное время контакта, очень медленное время контакта, быстрое время контакта и очень быстрое время контакта. Во время каждого условия метроном использовался для поддержания частоты шагов участников на уровне, соответствующем их самостоятельно выбранной частоте. Конкретные словесные инструкции заключались в том, чтобы «установить контакт с землей в такт ударам метронома и отреагировать на предоставленный сигнал.Вербальный сигнал подавался каждые 30 с (Moore et al., 2019) и заключался в следующем: условие (1) увеличить время контакта больше, чем обычно; условие (2) максимально увеличить время контакта; условие (3) уменьшить время контакта больше, чем обычно и; условие (4) максимально уменьшить время контакта. На протяжении каждого цикла дыхательные данные регистрировались с использованием онлайн-системы газового анализа (OxyconPro, Jaeger в Viasys Healthcare, Уорик, Великобритания), а RPE регистрировался по шкале Борга 6–20 (Borg, 1998) в конце каждого цикла. бегать.Все участники были одеты в свою обычную тренировочную форму и бегали в собственных кроссовках.
На протяжении каждого цикла дыхательные данные регистрировались с использованием онлайн-системы газового анализа (OxyconPro, Jaeger в Viasys Healthcare, Уорик, Великобритания), а RPE регистрировался по шкале Борга 6–20 (Borg, 1998) в конце каждого цикла. бегать.Все участники были одеты в свою обычную тренировочную форму и бегали в собственных кроссовках.
Сбор данных и расчет
Участников записали на видео во фронтальной плоскости с помощью приложения Runmatic (250 Гц) на iPhone, чтобы можно было определить время контакта левой и правой ноги с землей. Установка соответствовала предыдущим рекомендациям, согласно которым iPhone держится на расстоянии 30 см от задней части беговой дорожки, вертикально на одной высоте с беговой дорожкой (Balsalobre-Fernández et al., 2017). Было показано, что приложение Runmatic обеспечивает достоверное измерение времени контакта с землей (ICC > 0,96 при измерении критерия) и высокую надежность в течение сеанса при использовании 10-футовых контактов (α = 0,996) (Balsalobre-Fernández et al. , 2017). ). 10-секундная запись была сделана в течение 4-й минуты каждого состояния, что привело к пяти циклам походки (10 стоп-контактов). Видео были оцифрованы вручную в приложении Runmatic по одному и тому же индивидуальному, и данные о времени контакта с землей, времени полета и частоте шагов были экспортированы для каждого условия.Время контакта с землей (с) определяли как время между первоначальным контактом стопы и отрывом пальцев одной и той же стопы, в то время как время полета (с) представляло собой время между отрывом стопы одной ноги и первоначальным контактом другой стопы. Наконец, частота шагов (Гц) представляет собой количество касаний стопы (левой и правой) в течение одной единицы времени (с). Следуя рекомендациям Morin et al. (2005), жесткость ног (Н · м −1 ) была рассчитана с использованием экспортированного времени контакта с землей и полета, расчетной пиковой вертикальной силы (Н) методом синусоидальной волны и смоделированного вертикального смещения центра масс ( м) при контакте с землей.
, 2017). ). 10-секундная запись была сделана в течение 4-й минуты каждого состояния, что привело к пяти циклам походки (10 стоп-контактов). Видео были оцифрованы вручную в приложении Runmatic по одному и тому же индивидуальному, и данные о времени контакта с землей, времени полета и частоте шагов были экспортированы для каждого условия.Время контакта с землей (с) определяли как время между первоначальным контактом стопы и отрывом пальцев одной и той же стопы, в то время как время полета (с) представляло собой время между отрывом стопы одной ноги и первоначальным контактом другой стопы. Наконец, частота шагов (Гц) представляет собой количество касаний стопы (левой и правой) в течение одной единицы времени (с). Следуя рекомендациям Morin et al. (2005), жесткость ног (Н · м −1 ) была рассчитана с использованием экспортированного времени контакта с землей и полета, расчетной пиковой вертикальной силы (Н) методом синусоидальной волны и смоделированного вертикального смещения центра масс ( м) при контакте с землей. Полную информацию можно найти в Дополнении 1. Изменение единицы отношения между временем контакта с землей и жесткостью ног было рассчитано с использованием градиента уклона, созданного интерполированным временем контакта с землей и интерполированной жесткостью ног. Процедура интерполяции описана ниже.
Полную информацию можно найти в Дополнении 1. Изменение единицы отношения между временем контакта с землей и жесткостью ног было рассчитано с использованием градиента уклона, созданного интерполированным временем контакта с землей и интерполированной жесткостью ног. Процедура интерполяции описана ниже.
Данные о потреблении кислорода были отфильтрованы с использованием рекурсивного низкочастотного фильтра Баттерворта второго порядка (частота среза 0,43 Гц, определенная с помощью остаточного анализа), а средние метаболические затраты были рассчитаны с использованием последних 2 минут каждого запуска.Наборы данных были проверены на наличие выбросов (на 2 SD от среднего значения), и любые выбросы внутри участников для каждого запуска были удалены до расчета средней метаболической стоимости. Все данные о потреблении кислорода были визуально проверены на наличие стационарного состояния. Метаболические затраты рассчитывали с использованием кислорода, потребляемого на единицу массы тела в единицу времени (мл O 2 · кг -1 · мин -1 ). Использование метаболических затрат на единицу расстояния (мл O 2 · кг -1 · км -1 ), а не на единицу времени, не изменило отношения, выявленные в исследовании.
Использование метаболических затрат на единицу расстояния (мл O 2 · кг -1 · км -1 ), а не на единицу времени, не изменило отношения, выявленные в исследовании.
Оптимальное время контакта с землей определялось отдельно для каждого участника с использованием метаболических затрат. В частности, была рассчитана кубическая интерполяция по методу наименьших квадратов (многочлен третьего порядка, интерполированный до пятидесяти точек данных) со временем контакта с землей в качестве независимой переменной и метаболическими затратами в качестве зависимой переменной. Кубическая интерполяция использовалась для учета потенциального асимметричного увеличения метаболических затрат по обе стороны от оптимума и любого асимметричного увеличения и уменьшения контакта с землей, поскольку величины изменений времени контакта с землей нельзя было контролировать.Кубическая интерполяция была ограничена обычным временем контакта с землей и потреблением кислорода, являющимся известной фиксированной точкой полинома третьего порядка. Минимум кубической интерполяции был определен с использованием функции fmincon в MATLAB (Mathworks, Inc., 2018b) между следующими границами: самое быстрое время контакта с землей (нижняя граница) и самое медленное время контакта с землей (верхняя граница). Процедура была повторена для жесткости ног в качестве независимой переменной и метаболических затрат в качестве зависимой переменной.На рис. 1 показан пример измеренных и интерполированных данных. Все расчеты проводились в среде MATLAB. Было разработано бесплатно загружаемое программное обеспечение, позволяющее другим пользователям вычислять оптимальные характеристики походки (Moore, 2019).
Минимум кубической интерполяции был определен с использованием функции fmincon в MATLAB (Mathworks, Inc., 2018b) между следующими границами: самое быстрое время контакта с землей (нижняя граница) и самое медленное время контакта с землей (верхняя граница). Процедура была повторена для жесткости ног в качестве независимой переменной и метаболических затрат в качестве зависимой переменной.На рис. 1 показан пример измеренных и интерполированных данных. Все расчеты проводились в среде MATLAB. Было разработано бесплатно загружаемое программное обеспечение, позволяющее другим пользователям вычислять оптимальные характеристики походки (Moore, 2019).
Рисунок 1 . Пример измеренных (закрашенные кружки) и интерполированных (пунктирная линия) данных, показывающих взаимосвязь между отклонениями от самостоятельно выбранных характеристик беговой походки (%) и метаболическими затратами при самостоятельно выбранной походке (%). (A) Время контакта с землей (• и черная пунктирная линия). (B) Тугоподвижность ног ( и красная пунктирная линия). Оба соотношения показаны в одном масштабе по осям x и y, чтобы подчеркнуть различия в крутизне склона в основании кривой (окружающий минимум).
(B) Тугоподвижность ног ( и красная пунктирная линия). Оба соотношения показаны в одном масштабе по осям x и y, чтобы подчеркнуть различия в крутизне склона в основании кривой (окружающий минимум).
Статистический анализ
Средние значения (SD) биомеханических переменных, полученных как влево, так и вправо, были рассчитаны для каждого человека в каждом состоянии. После проверки нормальности с использованием Шарипо-Уилка ( W = 0,980, p = 0.551) был использован однофакторный дисперсионный анализ с повторными измерениями, чтобы проверить, сохраняется ли частота шагов во всех условиях бега. Поскольку данные RPE являются порядковыми, ранговый тест Спирмена использовался для оценки связи между RPE и метаболическими затратами, а частичный ранговый тест Спирмена использовался для оценки той же связи, но с контролируемым типом условий бега (привычным и управляемым). Малая, средняя и большая сила ассоциации была определена как 0,10–0,29, 0,30–0,49 и ≥0,5 соответственно. Статистический анализ проводился с помощью RStudio (версия 1.1.456, Бостон, Массачусетс) с альфа-каналом, равным 0,05.
Статистический анализ проводился с помощью RStudio (версия 1.1.456, Бостон, Массачусетс) с альфа-каналом, равным 0,05.
Результаты
Шаговая частота [ F (1, 40) = 0,051, p = 0,995] оказалась одинаковой во всех условиях (среднее значение ± стандартное отклонение: 2,67 ± 0,15 Гц). Это подтверждает, что привычная частота шагов сохранялась на всем протяжении. Среднее самостоятельно выбранное время контакта с землей составило 0,247 ± 0,016 с со средними метаболическими затратами 45,34 ± 5,42 мл O 2 · кг -1 · мин -1 Математическое оптимальное время контакта с землей было определено для всех участников, использующих полином третьего порядка, с большой долей дисперсии в стоимости метаболизма, объясняемой временем контакта с землей ( r 2 = 0.840; Таблица 1). На индивидуальном уровне шесть участников использовали самостоятельно выбранное время контакта с землей, которое было на 1–8 % короче их математического оптимума, в то время как остальные четыре участника использовали самостоятельно выбранное время контакта с землей, которое было на 1–5 % больше их математического оптимума. оптимальный (табл. 1; рис. 1).
оптимальный (табл. 1; рис. 1).
Таблица 1 . Самостоятельно выбранные и математически оптимальные (% от самостоятельно выбранных) время контакта с землей и метаболические затраты для каждого участника с полиномиальной моделью третьего порядка ( r 2 ).
Средняя самостоятельно выбранная жесткость ноги составила 8,38 ± 1,33 кН · м -1 , при этом математически оптимальная жесткость ноги была определена для всех участников. Подобная величина дисперсии метаболических затрат может быть объяснена жесткостью ног ( r 2 = 0,826), как и временем контакта с землей. Большинство ( n = 7) участников использовали самостоятельно выбранную жесткость ног, которая была на 1–16% выше их математического оптимума по сравнению с тремя участниками, которые использовали самостоятельно выбранную жесткость ног, которая была на 1–6% ниже, чем их математический оптимум. их математический оптимум (табл. 1).
Половина участников ( n = 5) находились в пределах 1% от их оптимальных метаболических затрат, а все участники, кроме одного, находились в пределах 5% от их оптимальных метаболических затрат (рис.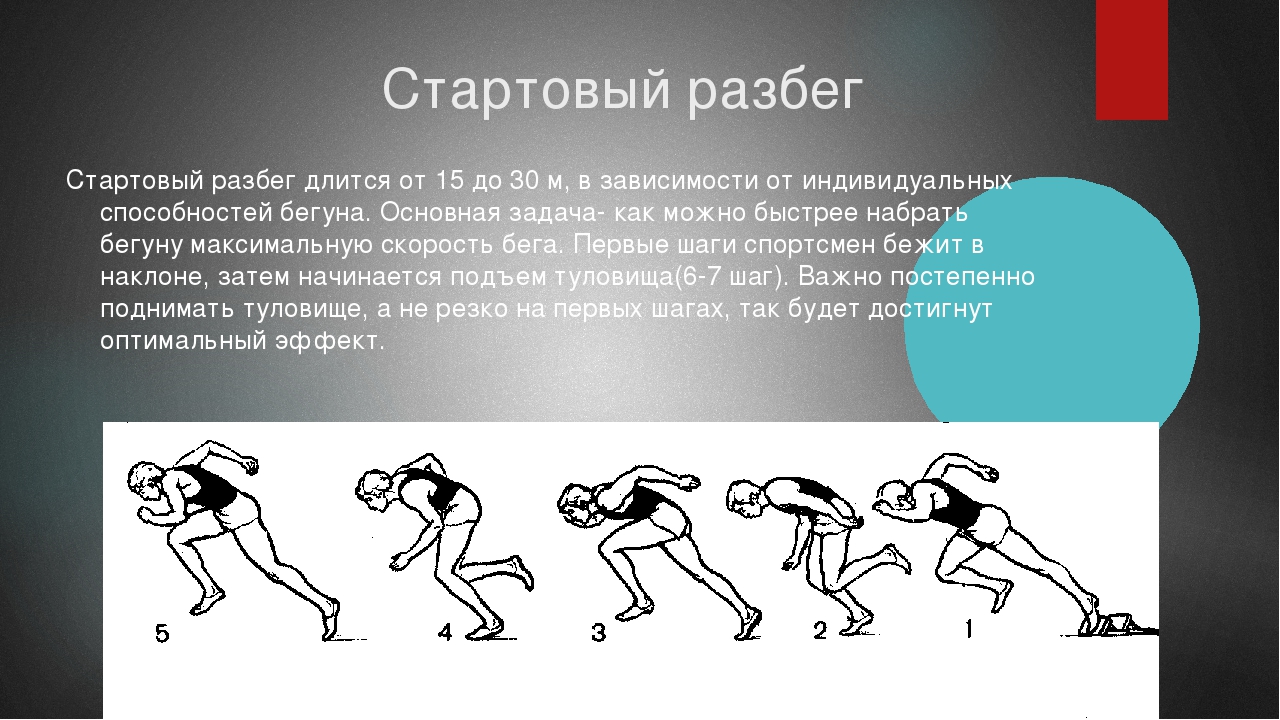 2). Примеры взаимосвязей между метаболическими затратами и временем контакта с землей представлены на рисунке 3 для трех различных реакций участников: время контакта с землей короче, близко к и дольше их математического оптимума. Также показаны соответствующие метаболические затраты и отношения жесткости ног.Среднее единичное изменение времени контакта с землей внутри участника по отношению к среднему единичному изменению жесткости ног составило 1:2,2 ± 0,2, что означает, что на каждый 1% изменения времени контакта с землей наблюдалось изменение жесткости ног на 2,2% (рис. 4). .
2). Примеры взаимосвязей между метаболическими затратами и временем контакта с землей представлены на рисунке 3 для трех различных реакций участников: время контакта с землей короче, близко к и дольше их математического оптимума. Также показаны соответствующие метаболические затраты и отношения жесткости ног.Среднее единичное изменение времени контакта с землей внутри участника по отношению к среднему единичному изменению жесткости ног составило 1:2,2 ± 0,2, что означает, что на каждый 1% изменения времени контакта с землей наблюдалось изменение жесткости ног на 2,2% (рис. 4). .
Рисунок 2 . Математически оптимальное время контакта с землей [мс; (A) ] и жесткость опоры [Н·м -1 ; (B) ] как отклонение от самостоятельно выбранных характеристик походки и соответствующее им улучшение метаболических затрат (мл.кг -1 · мин -1 ). Улучшение представляет собой снижение метаболических затрат по сравнению с метаболическими затратами, связанными с самостоятельно выбранной походкой. Черные точки (•) обозначают бегунов с более коротким, самостоятельно выбранным временем контакта с землей, чем оптимальное, и с более высокой жесткостью ног. Красные точки () представляют бегунов с более длительным временем контакта, чем оптимальное, и жесткостью голени.
Черные точки (•) обозначают бегунов с более коротким, самостоятельно выбранным временем контакта с землей, чем оптимальное, и с более высокой жесткостью ног. Красные точки () представляют бегунов с более длительным временем контакта, чем оптимальное, и жесткостью голени.
Рисунок 3 . Соотношение между отклонениями от самостоятельно выбранных характеристик беговой походки (%) и от метаболических затрат при самостоятельно выбранной походке (%). (A) Пример самостоятельно выбранного времени контакта с землей дольше оптимального. (B) Пример самостоятельно выбранного времени контакта с землей в пределах 1% от оптимального. (C) Пример самостоятельно выбранного времени контакта с землей меньше оптимального. Сплошные черные линии представляют время контакта с землей. Сплошные красные линии представляют жесткость ног. Оптимальные характеристики походки, минимизирующие метаболические затраты, обозначены кружками [черный (•) = время контакта с землей; красный () = жесткость ног]. Пунктирными линиями выделены соответствующие значения X и Y для оптимальных характеристик походки.
Пунктирными линиями выделены соответствующие значения X и Y для оптимальных характеристик походки.
Рисунок 4 . Среднее изменение времени контакта с землей и жесткости ног для группы () и каждого участника (). Пунктирная линия представляет собой линию наилучшего соответствия [жесткость ноги = 1,9 (время контакта с землей) + 0,12].
Наблюдалась значимая связь средней силы между RPE и метаболическими затратами ( r s = 0,358, p = 0,011). Когда привычное беговое состояние контролировалось с помощью частичной корреляции, сила ассоциации ослабевала ( r s = 0.302, р = 0,035).
Обсуждение
Целью этого исследования было изучить влияние манипулирования временем контакта с землей на метаболические затраты при беге, а также определить влияние измененной жесткости ног на метаболические затраты при беге. Это было первое исследование, в котором было установлено, что время контакта с землей и жесткость ног являются самооптимизируемыми характеристиками походки, поскольку было замечено, что тренированные бегуны работают на уровне своего математического экономического оптимума или близки к нему во время субмаксимального бега. Кроме того, почти все бегуны (90%) самостоятельно выбирали время контакта с землей и жесткость ног, которые производили метаболические затраты в пределах 5% от их математически оптимальных метаболических затрат. Эти результаты основаны на ранних работах Хогберга (1952) и Кавана и Уильямса (1982), предполагающих, что физиологическая и механическая адаптация, возникающая при многократном воздействии раздражителей, позволяет бегунам точно настраивать свою походку, чтобы минимизировать метаболические затраты при беге. Что касается метаболических затрат и RPE, хотя между переменными и существовала взаимосвязь, она ослабевала, когда контролировалось привычное состояние бега.Это говорит о том, что RPE не может быть полезным суррогатным показателем метаболических затрат при манипулировании походкой.
Кроме того, почти все бегуны (90%) самостоятельно выбирали время контакта с землей и жесткость ног, которые производили метаболические затраты в пределах 5% от их математически оптимальных метаболических затрат. Эти результаты основаны на ранних работах Хогберга (1952) и Кавана и Уильямса (1982), предполагающих, что физиологическая и механическая адаптация, возникающая при многократном воздействии раздражителей, позволяет бегунам точно настраивать свою походку, чтобы минимизировать метаболические затраты при беге. Что касается метаболических затрат и RPE, хотя между переменными и существовала взаимосвязь, она ослабевала, когда контролировалось привычное состояние бега.Это говорит о том, что RPE не может быть полезным суррогатным показателем метаболических затрат при манипулировании походкой.
В поддержку нашей первой гипотезы для всех бегунов наблюдался идентифицируемый оптимум (минимум), а также криволинейная U-образная зависимость между затратами на обмен веществ, временем контакта с землей и жесткостью ног (пример данных, рис.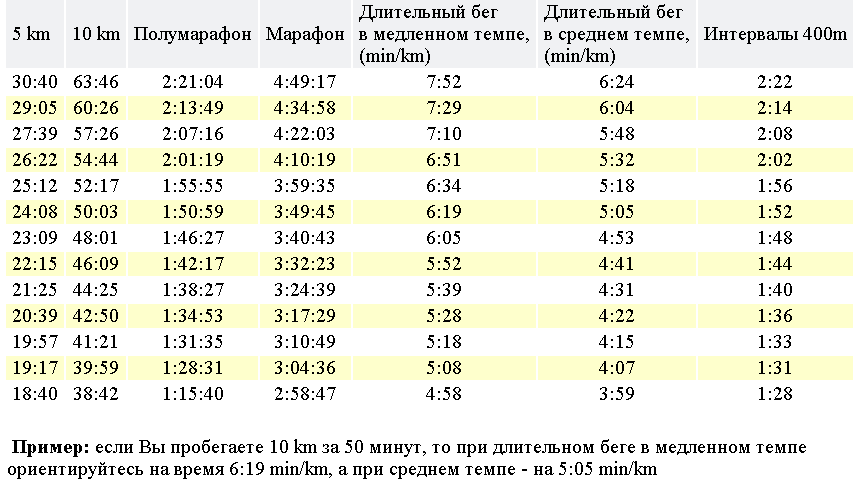 2). Эти результаты подтверждают теорию самооптимизации, но противоречат предыдущим исследованиям, которые утверждали, что они короче (Nummela et al., 2007; Santos-Concejero et al., 2014, 2017; Folland et al., 2017) или дольше (Williams and Cavanagh, 1986; Kram and Taylor, 1990; Di Michele and Merni, 2013) время контакта с землей является характеристикой экономичного бега, а большая жесткость ног снижает метаболические затраты (Dalleau et al., 1998). Однако в этих предыдущих исследованиях использовались перекрестные сравнения для определения различий в походке между бегунами. Такой подход обеспечивает богатую базу данных характеристик походки, которая может способствовать снижению метаболических затрат, однако индивидуальное профилирование реакции бегунов на манипуляции с походкой кажется более информативным для понимания экономичного бега.Таким образом, экстраполяция времени контакта с землей от одного бегуна к другому для оценки экономичного бега должна проводиться с осторожностью.
2). Эти результаты подтверждают теорию самооптимизации, но противоречат предыдущим исследованиям, которые утверждали, что они короче (Nummela et al., 2007; Santos-Concejero et al., 2014, 2017; Folland et al., 2017) или дольше (Williams and Cavanagh, 1986; Kram and Taylor, 1990; Di Michele and Merni, 2013) время контакта с землей является характеристикой экономичного бега, а большая жесткость ног снижает метаболические затраты (Dalleau et al., 1998). Однако в этих предыдущих исследованиях использовались перекрестные сравнения для определения различий в походке между бегунами. Такой подход обеспечивает богатую базу данных характеристик походки, которая может способствовать снижению метаболических затрат, однако индивидуальное профилирование реакции бегунов на манипуляции с походкой кажется более информативным для понимания экономичного бега.Таким образом, экстраполяция времени контакта с землей от одного бегуна к другому для оценки экономичного бега должна проводиться с осторожностью.
Время контакта с землей, по-видимому, имеет узкий оптимальный диапазон, в котором бегуны могут работать, вызывая изменения метаболических затрат при лишь незначительных изменениях времени контакта с землей, о чем свидетельствует относительно более крутой участок в основании кривой (окружающий минимум), чем нога. жесткость (рис. 1, 3). Напротив, жесткость ног имеет более плоские участки в основании кривых, черта, которая также является общей для длины и частоты шагов (Cavanagh and Williams, 1982).Такая черта может учитывать естественные вариации походки бегуна или отражать адаптацию к различным тренировочным стимулам, например, рельефу местности, скорости. Например, жесткость ног служит для поддержания стабильной беговой походки у людей и животных (Seyfarth et al., 2002) и может быстро регулироваться при беге по различным поверхностям (Ferris et al., 1998) и препятствиям (Birn-Jeffery et al., 1998). al., 2014) для сохранения смещения центра масс. Следовательно, возможность изменять жесткость ног в более широком оптимальном диапазоне, чем время контакта с землей, без увеличения метаболических затрат может быть выгодной экономической стратегией.Тем не менее, это также предполагает, что оптимальное время контакта с землей имеет большее значение, чем жесткость ног или частота/длина шага для экономически оптимальных критериев движения.
жесткость (рис. 1, 3). Напротив, жесткость ног имеет более плоские участки в основании кривых, черта, которая также является общей для длины и частоты шагов (Cavanagh and Williams, 1982).Такая черта может учитывать естественные вариации походки бегуна или отражать адаптацию к различным тренировочным стимулам, например, рельефу местности, скорости. Например, жесткость ног служит для поддержания стабильной беговой походки у людей и животных (Seyfarth et al., 2002) и может быстро регулироваться при беге по различным поверхностям (Ferris et al., 1998) и препятствиям (Birn-Jeffery et al., 1998). al., 2014) для сохранения смещения центра масс. Следовательно, возможность изменять жесткость ног в более широком оптимальном диапазоне, чем время контакта с землей, без увеличения метаболических затрат может быть выгодной экономической стратегией.Тем не менее, это также предполагает, что оптимальное время контакта с землей имеет большее значение, чем жесткость ног или частота/длина шага для экономически оптимальных критериев движения.
Во время стабильного бега с различной частотой шагов регулировка жесткости ног также приводит к созданию постоянной силы ног (Farley and Gonzalez, 1996; Seyfarth et al., 2002). Мы провели последующий корреляционный анализ, чтобы проверить, присутствует ли постоянное усилие на ноги, и обнаружили, что это не так, когда жесткость ног быстро регулируется, чтобы приспособиться к более коротким и более длительным временам контакта с землей.В частности, жесткость ног была положительно связана с силой ног (оценочная пиковая вертикальная сила; r с = 0,639, p < 0,001), тогда как если бы сила ног была постоянной, никакой связи не было бы. Ограничения скорости бега и частоты шагов, налагаемые на бегунов в текущем исследовании, ограничивали бы степень свободы, которую каждый бегун имел для регулировки жесткости своих ног, что потенциально приводило к этой очевидной однородной реакции измененной силы ног.Возможно, это отражает оптимизированную адаптацию, развившуюся в результате воздействия беговых и тренировочных стимулов, позволяющую тренированным бегунам быстро приспосабливаться к регулировке жесткости ног. Возможно, подобные гомогенные реакции не могут быть обнаружены у нетренированных бегунов, поскольку они демонстрируют менее последовательную реакцию на увеличение скорости бега и находятся дальше от своего математического оптимума, чем тренированные бегуны (de Ruiter et al., 2013; Bitchell et al., 2019), однако необходима дальнейшая работа в этом направлении.
Возможно, подобные гомогенные реакции не могут быть обнаружены у нетренированных бегунов, поскольку они демонстрируют менее последовательную реакцию на увеличение скорости бега и находятся дальше от своего математического оптимума, чем тренированные бегуны (de Ruiter et al., 2013; Bitchell et al., 2019), однако необходима дальнейшая работа в этом направлении.
Большинство ( n = 6) бегунов самостоятельно выбрали время контакта с землей и жесткость ног, которые были короче и выше, соответственно, их математически оптимальных значений. Это означает, что они способствуют производству быстрой и высокой вертикальной силы, создаваемой жесткой нижней конечностью. Объединив это понимание с предыдущей работой, которая выявила, что большинство тренированных бегунов предпочитали чрезмерный шаг (длительное время шага, чем оптимальное) (Cavanagh and Williams, 1982; de Ruiter et al., 2013), предполагает, что тренированные бегуны предпочитают низкий коэффициент нагрузки, что указывает на то, что они полагаются на накопление и высвобождение упругой энергии для минимизации метаболических затрат (Lussiana et al. , 2019). Чтобы достичь этого, мышцы также должны работать с более высокими скоростями сокращения, что потребует привлечения большего количества двигательных единиц для создания необходимых высоких усилий (Fletcher and MacIntosh, 2017). Такие факторы, как тренировочные стимулы и внутренние свойства мышц и сухожилий, могут означать, что костно-мышечная система бегуна настроена на такие требования.Однако такая механическая стратегия будет вызывать высокие скорости вертикальной и горизонтальной нагрузки и величины сил, связанных с ударом, что может подвергнуть бегуна риску травмы нижних конечностей (Hreljac et al., 2000; Napier et al., 2018). Несколько ( n = 4) бегунов использовали другую механическую стратегию, в результате чего у них было более длительное время контакта с землей и более податливая нога (менее жесткая), чем их математический оптимум. Это привело бы к более высокому коэффициенту нагрузки, чем оптимальный, что указывало бы на то, что бегуны отдавали приоритет горизонтальному смещению и уменьшали вертикальное смещение (Lussiana et al.
, 2019). Чтобы достичь этого, мышцы также должны работать с более высокими скоростями сокращения, что потребует привлечения большего количества двигательных единиц для создания необходимых высоких усилий (Fletcher and MacIntosh, 2017). Такие факторы, как тренировочные стимулы и внутренние свойства мышц и сухожилий, могут означать, что костно-мышечная система бегуна настроена на такие требования.Однако такая механическая стратегия будет вызывать высокие скорости вертикальной и горизонтальной нагрузки и величины сил, связанных с ударом, что может подвергнуть бегуна риску травмы нижних конечностей (Hreljac et al., 2000; Napier et al., 2018). Несколько ( n = 4) бегунов использовали другую механическую стратегию, в результате чего у них было более длительное время контакта с землей и более податливая нога (менее жесткая), чем их математический оптимум. Это привело бы к более высокому коэффициенту нагрузки, чем оптимальный, что указывало бы на то, что бегуны отдавали приоритет горизонтальному смещению и уменьшали вертикальное смещение (Lussiana et al. , 2019). В отличие от стратегии с низким коэффициентом нагрузки, мышцы будут работать с более низкой скоростью сокращения, и для создания меньшей силы потребуется задействовать меньше двигательных единиц (Fletcher and MacIntosh, 2017). Эта стратегия может указывать на недостаточную внутреннюю мышечно-сухожильную жесткость или на то, что приоритетом является сокращение работы против гравитации и сил, связанных с ударами.
, 2019). В отличие от стратегии с низким коэффициентом нагрузки, мышцы будут работать с более низкой скоростью сокращения, и для создания меньшей силы потребуется задействовать меньше двигательных единиц (Fletcher and MacIntosh, 2017). Эта стратегия может указывать на недостаточную внутреннюю мышечно-сухожильную жесткость или на то, что приоритетом является сокращение работы против гравитации и сил, связанных с ударами.
Поскольку выбор походки и самооптимизация считаются подсознательными процессами (Cavanagh and Williams, 1982; Moore et al., 2012), вполне возможно, что большинство бегунов неосознанно отдают приоритет минимизации метаболических затрат, а не минимизации потенциально вредных последствий, связанных с воздействием. сил.Это может быть связано с тем, что определение связанных с ударом сил скелетно-мышечной и нервной системами может быть не таким чувствительным, как определение метаболических потребностей сердечно-сосудистой системой. Даже при снятой обуви и, следовательно, усилении соматосенсорной обратной связи чувствительность подошвенной поверхности стопы не связана с пиковым давлением стопы во время фазы торможения (Nurse and Nigg, 1999). Учитывая, что соматосенсорная обратная связь потенциально может быть дополнительно ослаблена амортизацией, присутствующей в традиционной беговой обуви, неудивительно, что люди кажутся более приспособленными к метаболическим потребностям, а не к силе удара.
Учитывая, что соматосенсорная обратная связь потенциально может быть дополнительно ослаблена амортизацией, присутствующей в традиционной беговой обуви, неудивительно, что люди кажутся более приспособленными к метаболическим потребностям, а не к силе удара.
Проинструктировав бегунов сократить или увеличить время контакта с землей, мы смогли уникальным образом протестировать влияние времени контакта с землей на метаболические затраты при ограничении скорости бега, частоты/длины шагов и, таким образом, частоты/длины шага. Эти ограничения были важны, поскольку они, как известно, влияют на метаболические затраты (Gutmann et al., 2006). Интересно, что скорости бега и частоте/длине шага уделялось больше внимания, чем времени контакта с землей, которое в значительной степени игнорировалось во время ограниченного оптимизационного тестирования, когда манипулировали характеристиками походки (Knuttgen, 1961; Cavanagh and Williams, 1982; Gutmann et al., 2006; Хантер и Смит, 2007 г.; де Руитер и др. , 2013). Учитывая значительную роль времени контакта с землей в определении метаболических затрат у людей, а также у двуногих и четвероногих видов при ходьбе и беге (Taylor et al., 1980, 1982; Kram and Taylor, 1990; Roberts et al., 1998). ), эта оплошность могла привести к упрощению оптимизации передвижения. Кроме того, Флетчер и Макинтош (2017) утверждали, что бегуны сохраняют время контакта с землей, а не максимизируют накопление и отдачу упругой энергии из-за выбора более низкой частоты шага, чем оптимальная.Тем не менее, потребовав от скелетно-мышечной системы быстрой корректировки времени контакта с землей в стесненных условиях, мы смогли определить, что большинство бегунов, по-видимому, уделяют первостепенное внимание накоплению и высвобождению упругой энергии, что усилило необходимость учитывать время контакта с землей при передвижении. уравнение оптимизации.
, 2013). Учитывая значительную роль времени контакта с землей в определении метаболических затрат у людей, а также у двуногих и четвероногих видов при ходьбе и беге (Taylor et al., 1980, 1982; Kram and Taylor, 1990; Roberts et al., 1998). ), эта оплошность могла привести к упрощению оптимизации передвижения. Кроме того, Флетчер и Макинтош (2017) утверждали, что бегуны сохраняют время контакта с землей, а не максимизируют накопление и отдачу упругой энергии из-за выбора более низкой частоты шага, чем оптимальная.Тем не менее, потребовав от скелетно-мышечной системы быстрой корректировки времени контакта с землей в стесненных условиях, мы смогли определить, что большинство бегунов, по-видимому, уделяют первостепенное внимание накоплению и высвобождению упругой энергии, что усилило необходимость учитывать время контакта с землей при передвижении. уравнение оптимизации.
На сегодняшний день только одно исследование изменило характеристики походки в сторону математического оптимума человека. Трехнедельное вмешательство успешно изменило частоту шагов в сторону математического оптимума и снизило метаболические затраты у трех бегунов (Morgan et al., 1994), показывая полезность повторного обучения походке для ускорения процесса самооптимизации. Хотя для подтверждения этих выводов необходимы более масштабные исследования, мероприятия по биомеханической переподготовке, ориентированные на травмы, с участием более крупных групп показали, что желаемые изменения беговой походки могут быть достигнуты за аналогичный период времени (Crowell and Davis, 2011; Roper et al., 2016). Вмешательства, основанные на силе, также могут быть эффективными, но, вероятно, потребуют больше времени для физиологической адаптации. Например, время контакта с землей увеличилось на 3-4% после 8- (Ferrauti et al., 2010) и 12-недельное (Giovanelli et al., 2017) силовое вмешательство. Интересно, что плиометрическая тренировка, которую часто рекомендуют бегунам, поскольку она направлена на улучшение цикла растяжения-сокращения и характеристик жесткости человека, не имеет доказательств того, что короткое время контакта с землей, поощряемое во время тренировки, переносится на беговую походку (Giovanelli et al.
Трехнедельное вмешательство успешно изменило частоту шагов в сторону математического оптимума и снизило метаболические затраты у трех бегунов (Morgan et al., 1994), показывая полезность повторного обучения походке для ускорения процесса самооптимизации. Хотя для подтверждения этих выводов необходимы более масштабные исследования, мероприятия по биомеханической переподготовке, ориентированные на травмы, с участием более крупных групп показали, что желаемые изменения беговой походки могут быть достигнуты за аналогичный период времени (Crowell and Davis, 2011; Roper et al., 2016). Вмешательства, основанные на силе, также могут быть эффективными, но, вероятно, потребуют больше времени для физиологической адаптации. Например, время контакта с землей увеличилось на 3-4% после 8- (Ferrauti et al., 2010) и 12-недельное (Giovanelli et al., 2017) силовое вмешательство. Интересно, что плиометрическая тренировка, которую часто рекомендуют бегунам, поскольку она направлена на улучшение цикла растяжения-сокращения и характеристик жесткости человека, не имеет доказательств того, что короткое время контакта с землей, поощряемое во время тренировки, переносится на беговую походку (Giovanelli et al. и др., 2017; Гомес-Молина и др., 2018). Наше исследование показывает, что тренированные бегуны способны изменять жесткость ног, следуя биомеханическим инструкциям и среднему коэффициенту изменения единицы (1:2.2) подтверждает предыдущие сообщения о том, что 5-процентное изменение времени контакта с землей соответствует приблизительно 10-процентному изменению жесткости ног (Морин и др., 2005, 2007). Таким образом, биомеханическая переподготовка рекомендуется в качестве первого подхода к вмешательству, если изменения жесткости являются целевыми из-за более коротких требований по времени и возможности непрерывной переоценки времени контакта с землей во время каждой тренировочной сессии.
и др., 2017; Гомес-Молина и др., 2018). Наше исследование показывает, что тренированные бегуны способны изменять жесткость ног, следуя биомеханическим инструкциям и среднему коэффициенту изменения единицы (1:2.2) подтверждает предыдущие сообщения о том, что 5-процентное изменение времени контакта с землей соответствует приблизительно 10-процентному изменению жесткости ног (Морин и др., 2005, 2007). Таким образом, биомеханическая переподготовка рекомендуется в качестве первого подхода к вмешательству, если изменения жесткости являются целевыми из-за более коротких требований по времени и возможности непрерывной переоценки времени контакта с землей во время каждой тренировочной сессии.
Связь средней силы между воспринимаемыми усилиями и метаболическими затратами в текущем исследовании подтверждает нашу вторую гипотезу, но ниже критерия, представленного Chen et al.(2002) в своем мета-анализе упражнений на беговой дорожке (95% ДИ для r = 0,478–0,629) и субмаксимальных упражнений (95% ДИ для r = 0,766–0,870). Когда привычное состояние бега контролировалось для ослабления связи, можно предположить, что нарушенная походка привела к разрыву между метаболическими затратами и воспринимаемым усилием, как ранее наблюдалось в нашей лаборатории (Moore et al., 2019). Акт манипулирования беговой походкой с помощью словесных сигналов, вероятно, сместил фокус внимания и усилил ощущаемое усилие механического требования бега.Следовательно, на основе результатов исследования, в дополнение к недавней работе (Moore et al., 2019), использование воспринимаемого усилия в качестве суррогата для определения влияния изменения беговой походки на метаболические затраты и / или использование его для мониторинга тренировочных реакций, ориентированных на технику. из-за его связи с метаболическими затратами следует проводить с осторожностью.
Когда привычное состояние бега контролировалось для ослабления связи, можно предположить, что нарушенная походка привела к разрыву между метаболическими затратами и воспринимаемым усилием, как ранее наблюдалось в нашей лаборатории (Moore et al., 2019). Акт манипулирования беговой походкой с помощью словесных сигналов, вероятно, сместил фокус внимания и усилил ощущаемое усилие механического требования бега.Следовательно, на основе результатов исследования, в дополнение к недавней работе (Moore et al., 2019), использование воспринимаемого усилия в качестве суррогата для определения влияния изменения беговой походки на метаболические затраты и / или использование его для мониторинга тренировочных реакций, ориентированных на технику. из-за его связи с метаболическими затратами следует проводить с осторожностью.
Мы признаем, что в этом исследовании было несколько ограничений. Несмотря на то, что все участники получали одни и те же сигналы, индивидуальные интерпретации приводили к самостоятельным изменениям времени контакта с землей. Это привело к тому, что некоторые бегуны увеличивали и уменьшали время контакта с землей больше, чем другие. Хотя нам не удалось преодолеть это в нашей лаборатории, мы считаем, что такая подсказка представляет собой полезную стратегию переобучения походке для тренеров и практиков. Кроме того, из-за различий между участниками во времени манипулируемого контакта с землей анализ был сосредоточен на индивидуальных реакциях, а не на групповых отношениях. Однако этот подход позволяет выявить ряд реакций, которые тренеры и практикующие врачи также могут наблюдать и количественно оценивать с помощью разработанного бесплатного программного обеспечения (Moore, 2019).Жесткость ног оценивалась, а не измерялась с использованием методов золотого стандарта. Тем не менее, расчеты, которые были использованы, были проверены как для наземного бега, так и для беговой дорожки, показывая низкий уровень систематической ошибки (6%) для последнего (Morin et al., 2005). Кроме того, аналогичное взвешивание времени контакта с землей с жесткостью ног, определенное в этом исследовании, по сравнению с предыдущими экспериментальными и теоретическими данными, подтверждает предположение, что оно может отражать поведение человека при беге.
Это привело к тому, что некоторые бегуны увеличивали и уменьшали время контакта с землей больше, чем другие. Хотя нам не удалось преодолеть это в нашей лаборатории, мы считаем, что такая подсказка представляет собой полезную стратегию переобучения походке для тренеров и практиков. Кроме того, из-за различий между участниками во времени манипулируемого контакта с землей анализ был сосредоточен на индивидуальных реакциях, а не на групповых отношениях. Однако этот подход позволяет выявить ряд реакций, которые тренеры и практикующие врачи также могут наблюдать и количественно оценивать с помощью разработанного бесплатного программного обеспечения (Moore, 2019).Жесткость ног оценивалась, а не измерялась с использованием методов золотого стандарта. Тем не менее, расчеты, которые были использованы, были проверены как для наземного бега, так и для беговой дорожки, показывая низкий уровень систематической ошибки (6%) для последнего (Morin et al., 2005). Кроме того, аналогичное взвешивание времени контакта с землей с жесткостью ног, определенное в этом исследовании, по сравнению с предыдущими экспериментальными и теоретическими данными, подтверждает предположение, что оно может отражать поведение человека при беге.
Заключение
Было показано, что время контакта с землей и жесткость ног автоматически оптимизируются в группе тренированных бегунов, при этом все бегуны, кроме одного, находятся в пределах 5% от их оптимальных метаболических затрат во время их обычного бегового аллюра.Кроме того, для всех бегунов были обнаружены идентифицируемые минимумы, указывающие на наличие криволинейной, U-образной зависимости между затратами на метаболизм, временем контакта с землей и жесткостью ног. Бегуны работали в более узком диапазоне оптимального времени контакта с землей, чем жесткость ног, когда скорость бега и частота шагов были ограничены. Следовательно, оптимальное время контакта с землей может иметь большее значение для экономически оптимальных критериев движения, чем жесткость ног. Большинство бегунов предпочли немного более короткое время контакта с землей и более высокую жесткость ног, чем оптимально, что свидетельствует о зависимости от накопления и высвобождения упругой энергии и о том, что человеческое тело может быть настроено на минимизацию метаболических затрат, а не сил, связанных с ударом. Манипуляции с беговой походкой нарушают взаимосвязь между метаболическими затратами и воспринимаемыми усилиями, поэтому тренерам и практикующим врачам не рекомендуется использовать RPE в качестве суррогатного показателя при оценке экономичной беговой походки.
Манипуляции с беговой походкой нарушают взаимосвязь между метаболическими затратами и воспринимаемыми усилиями, поэтому тренерам и практикующим врачам не рекомендуется использовать RPE в качестве суррогатного показателя при оценке экономичной беговой походки.
Заявление о доступности данных
Наборы данных, созданные для этого исследования, можно найти на Figshare https://doi.org/10.25401/cardiffmet.8323307.v2.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены Кардиффской школой спорта и медицинских наук Кардиффского столичного университета.Пациенты/участники предоставили письменное информированное согласие на участие в этом исследовании.
Вклад авторов
ISM и KJA задумали и разработали исследование и составили рукопись. CC и JH набрали участников и провели сбор данных. MM-R помогал в разработке дизайна исследования и сборе данных. ИСМ провел расчетно-статистический анализ. HSRJ внес свой вклад в подготовку рукописи. Все авторы предоставили критическую информацию в окончательной версии.
Все авторы предоставили критическую информацию в окончательной версии.
Финансирование
IM получил финансирование от Британской ассоциации науки о спорте и физических упражнениях для получения награды для начинающих исследователей и практиков.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы поблагодарить Майкла Лонга за технические знания, которые он предоставил во время исследования.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fspor.2019.00053/full#supplementary-material
.Ссылки
Балсалобре-Фернандес, К., Агопян, Х., и Морен, Ж.-Б. (2017). Достоверность и надежность приложения для iPhone для измерения беговой механики. J. Appl. Биомех. 33, 222–226. doi: 10.1123/jab.2016-0104
J. Appl. Биомех. 33, 222–226. doi: 10.1123/jab.2016-0104
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бирн-Джеффери, А.В., Хубики С.М., Блюм Ю., Реневский Д., Херст Дж. В. и Дейли М. А. (2014). Не сломайте ногу: при перегоне птиц от перепела к страусу безопасность ног и экономичность на неровной местности имеют приоритетное значение. Дж. Экспл. биол. 217, 3786–3796. doi: 10.1242/jeb.102640
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Битчелл, К.Л., Маккарти-Райан, М., Гум, Т., и Мур, И.С. (2019). Пружинно-массовые характеристики при передвижении человека: опыт бега и физиологические аспекты накопления лактата в крови. евро. Дж. Спортивные науки. 19, 1328–1335. дои: 10.1080/17461391.2019.1609095
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Борг, Г. (1998). Воспринимаемое напряжение и боль Борга . Шампейн, Иллинойс: Кинетика человека.
Академия Google
Кавана, П.Р., и Уильямс, К.Р. (1982). Влияние изменения длины шага на потребление кислорода во время бега на длинные дистанции. Мед. науч. Спортивное упражнение. 14, 30–35. дои: 10.1249/00005768-198201000-00006
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чен, М.Дж., Фан, X., и Мо, С.Т. (2002). Связанная с критериями валидность оценок Борга по шкале воспринимаемой нагрузки у здоровых людей: метаанализ. J. Sports Sci. 20, 873–899. дои: 10.1080/026404102320761787
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Конник, М. Дж., и Ли, Ф. К. (2014). Изменение времени мышечных сокращений и экономичности бега с изменением шага во время бега. Осанка походки 39, 634–637. дои: 10.1016/j.gaitpost.2013.07.112
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Далло Г., Белли А., Бурден М. и Лакур Ж.-Р. (1998). Модель пружинной массы и энергозатраты на бег на беговой дорожке. евро. Дж. Заявл. Физиол. 77, 257–263. дои: 10.1007/s004210050330
Модель пружинной массы и энергозатраты на бег на беговой дорожке. евро. Дж. Заявл. Физиол. 77, 257–263. дои: 10.1007/s004210050330
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
де Рюйтер, С. Дж., Вердейк, П. В., Веркер, В., Зуидема, М. Дж., и де Хаан, А. (2013). Частота шагов в зависимости от потребления кислорода у опытных и начинающих бегунов. евро. Дж. Спортивные науки. 14, 251–258. дои: 10.1080/17461391.2013.783627
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ди Микеле, Р., и Мерни, Ф. (2013). Одновременное влияние модели удара и времени контакта с землей на экономичность бега. J. Sci. Мед. Спорт . 17, 414–418. doi: 10.1016/j.jsams.2013.05.012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ферраути А., Бергерманн М. и Фернандес-Фернандес Дж.(2010). Влияние одновременных тренировок на силу и выносливость на беговые характеристики и экономичность бега у марафонцев-любителей. Дж. Сила конд. Рез. 24, 2770–2778. doi: 10.1519/JSC.0b013e3181d64e9c
Дж. Сила конд. Рез. 24, 2770–2778. doi: 10.1519/JSC.0b013e3181d64e9c
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Феррис, Д. П., Луи, М., и Фарли, К. Т. (1998). Бег в реальном мире: регулировка жесткости ног для разных поверхностей. Проц. Р. Соц. В 265, 989–994. doi: 10.1098/rspb.1998.0388
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фолланд, Дж. П., Аллен, С. Дж., Блэк, М. И., Хэндсейкер, Дж. К., и Форрестер, С. Э. (2017). Техника бега является важным компонентом экономичности и производительности бега. Мед. науч. Спортивное упражнение. 49, 1412–1423. doi: 10.1249/MSS.0000000000001245
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джованелли Н., Табога П., Рейц Э. и Лаззер С. (2017). Влияние силовых, взрывных и плиометрических тренировок на энергозатраты при беге у спортсменов со сверхвыносливостью. евро. Дж. Спортивные науки. 17, 805–813. дои: 10.1080/17461391.2017.1305454
17, 805–813. дои: 10.1080/17461391.2017.1305454
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гомес-Молина, Дж., Огета-Алдай, А., Камара, Дж., Стикли, К., и Гарсия-Лопес, Дж. (2018). Влияние 8 недель одновременных плиометрических и беговых тренировок на пространственно-временные и физиологические параметры начинающих бегунов. евро. Дж. Спортивные науки. 18, 162–169. дои: 10.1080/17461391.2017.1404133
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хойт, Р.В., Кнапик, Дж. Дж., Ланза, Дж. Ф., Джонс, Б. Х., и Стааб, Дж. С. (1994). Амбулаторный монитор контакта стопы для оценки метаболических затрат на передвижение человека. J. Appl. Физиол . 76, 1818–1822 гг. doi: 10.1152/jappl.1994.76.4.1818
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Греляц, А., Маршалл, Р. Н., и Хьюм, П. А. (2000). Оценка потенциальной травмы от перенапряжения нижних конечностей у бегунов. Мед. науч. Спортивное упражнение. 32, 1635–1641. дои: 10.1097/00005768-200009000-00018
Мед. науч. Спортивное упражнение. 32, 1635–1641. дои: 10.1097/00005768-200009000-00018
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хантер, И., и Смит, Г.А. (2007). Предпочтительная и оптимальная частота шагов, жесткость и экономичность: изменения в зависимости от утомления во время высокоинтенсивного бега в течение 1 часа. евро. Дж. Заявл. Физиол. 100, 653–661. doi: 10.1007/s00421-007-0456-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кипп, С., Грабовски, А. М., и Крам, Р. (2018). Что определяет метаболические затраты человека на бег в широком диапазоне скоростей? Дж.Эксп. биол. 221:jeb184218. doi: 10.1242/jeb.184218
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Knuttgen, HG (1961). Потребление кислорода и частота пульса при беге с неопределенной и определенной длиной шага с разной скоростью. Acta Physiol. Сканд. 52, 366–371. doi: 10.1111/j.1748-1716.1961.tb02232.x
doi: 10.1111/j.1748-1716.1961.tb02232.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Луссиана, Т., Патоз, А., Жиндр, К., Муро, Л., и Эбер-Лозье, К.(2019). Влияние времени на земле на экономичность: меньше не всегда лучше. Дж. Экспл. биол. 222:jeb192047. doi: 10.1242/jeb.192047
Полнотекстовая перекрестная ссылка | Академия Google
McLaren, S.J., Macpherson, T.W., Coutts, A.J., Hurst, C., Spears, I.R., and Weston, M. (2018). Отношения между внутренними и внешними показателями тренировочной нагрузки и интенсивности в командных видах спорта: метаанализ. Спорт Мед. 48, 641–658. doi: 10.1007/s40279-017-0830-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мур, И.(2019). Программное обеспечение для определения смоделированных оптимальных характеристик походки (необходимо время выполнения). Фиг. . doi: 10.25401/cardiffmet.8323283
Полнотекстовая перекрестная ссылка | Академия Google
Мур, И. С. (2016). Существует ли экономичная техника бега? Обзор модифицируемых биомеханических факторов, влияющих на экономичность бега. Спорт Мед. 46, 793–807. doi: 10.1007/s40279-016-0474-4
С. (2016). Существует ли экономичная техника бега? Обзор модифицируемых биомеханических факторов, влияющих на экономичность бега. Спорт Мед. 46, 793–807. doi: 10.1007/s40279-016-0474-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мур, И.С., Джонс, А.М., и Диксон, С.Дж. (2012). Механизмы улучшения экономичности бега у начинающих бегунов. Мед. науч. Спортивное упражнение. 44, 1756–1763. doi: 10.1249/MSS.0b013e318255a727
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мур, И.С., Джонс, А.М., и Диксон, С.Дж. (2016). Снижение затрат кислорода при беге связано с выравниванием результирующего GRF и вектора оси ноги: пилотное исследование. Скан. Дж. Мед. науч. Спорт . 26, 809–815. дои: 10.1111/смс.12514
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мур, И. С., Филлипс, Д. Дж., Эшфорд, К. А., Маллен, Р., Гум, Т., и Гиттоуз, М. Р. Дж. (2019). Междисциплинарное исследование стратегий концентрации внимания, используемых во время повторного обучения ходьбе при беге. Scand J. Med. науч. Спорт 29, 1572–1582. doi: 10.1111/смс.13490
Междисциплинарное исследование стратегий концентрации внимания, используемых во время повторного обучения ходьбе при беге. Scand J. Med. науч. Спорт 29, 1572–1582. doi: 10.1111/смс.13490
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Морган, Д., Мартин, П., Крейб, М., Карузо, К., Клифтон Р. и Хоупвелл Р. (1994). Влияние оптимизации длины шага на аэробные потребности бега. J Appl Physiol . 77, 245–251. doi: 10.1152/jappl.1994.77.1.245
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Морин, Дж. Б., Далло Г. Фау — Киролайнен, Х., Киролайнен Х. Фау — Жаннин, Т., Жаннин Т. Фау — Белли, А., и Белли, А. (2005). Простой метод измерения жесткости во время бега. J. Appl. Биомех. 21, 167–180. дои: 10.1123/jab.21.2.167
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Морин, Дж. Б., Самозино, П., Замезиати, К., и Белли, А. (2007). Влияние измененной частоты шага и времени контакта на поведение ног при беге. Дж. Биомех. 40, 3341–3348. doi: 10.1016/j.jbiomech.2007.05.001
Дж. Биомех. 40, 3341–3348. doi: 10.1016/j.jbiomech.2007.05.001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нейпир, К., Маклин, К.Л., Маурер, Дж., Тонтон, Дж.Э., и Хант, Массачусетс (2018).Кинетические факторы риска беговых травм у бегунов-любителей. Скан. Дж. Мед. науч. Спорт . 28, 2164–2172. doi: 10.1111/смс.13228
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Медсестра, Массачусетс, и Нигг, Б.М. (1999). Количественная оценка взаимосвязи тактильной и вибрационной чувствительности стопы человека с распределением подошвенного давления во время ходьбы. Клин Биомех . 14, 667–672. дои: 10.1016/S0268-0033(99)00020-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Робертс, Т.Дж., Крам Р., Вейанд П.Г. и Тейлор С.Р. (1998). Энергетика двуногого бега. I. Метаболические затраты на создание силы. Дж. Экспл. Биол . 201, 2745–51.
Реферат PubMed | Академия Google
Ропер, Дж. Л., Хардинг, Э. М., Доерфлер, Д., Декстер, Дж. Г., Кравиц, Л., Дуфек, Дж. С., и соавт. (2016). Эффекты повторного обучения походке у бегунов с болью в надколеннике и бедренной кости: рандомизированное исследование. клин. Биомех . 35:14–22. doi: 10.1016/j.clinbiomech.2016.03.010
Л., Хардинг, Э. М., Доерфлер, Д., Декстер, Дж. Г., Кравиц, Л., Дуфек, Дж. С., и соавт. (2016). Эффекты повторного обучения походке у бегунов с болью в надколеннике и бедренной кости: рандомизированное исследование. клин. Биомех . 35:14–22. doi: 10.1016/j.clinbiomech.2016.03.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сантос-Консехеро, Дж., Там, Н., Кутзи, Д. Р., Оливан, Дж., Ноукс, Т. Д., и Такер, Р. (2017). Связаны ли характеристики походки и силы опорной реакции с затратами энергии при беге у элитных кенийских бегунов? J. Sports Sci. 35, 531–538. дои: 10.1080/02640414.2016.1175655
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сантос-Консехеро, Дж., Там, Н., Гранадос, К., Иразуста, Дж., Бидаурразага-Летона, И., Забала-Лили, Дж., и другие. (2014). Угол шага как новый показатель экономичности бега у хорошо тренированных бегунов. Дж. Сила конд. Рез. 28, 1889–1895 гг. doi: 10. 1519/JSC.0000000000000325
1519/JSC.0000000000000325
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тейлор, К., Хеглунд, Н., МакМахон, Т., и Луни, Т. (1980). Энергетические затраты на создание мышечной силы во время бега — сравнение крупных и мелких животных. Дж. Экспл. биол. 86, 9–18.
Академия Google
Taylor, C.R., Heglund, N.C., and Maloiy, G.M. (1982). Энергетика и механика земного передвижения.I. Потребление метаболической энергии в зависимости от скорости и размера тела у птиц и млекопитающих. Дж. Экспл. биол. 97, 1–21. doi: 10.1146/annurev.ph.44.030182.000525
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
van Oeveren, B.T., de Ruiter, CJ, Beek, PJ, and van Dien, JH (2017). Оптимальная частота шагов при беге с разной скоростью. PLoS ONE 12:e0184273. doi: 10.1371/journal.pone.0184273
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уильямс, К. Р. и Кавана П.Р. (1987). Взаимосвязь между механикой бега на длинные дистанции, экономичностью бега и результативностью. J Appl Physiol . 63, 1236–1245. doi: 10.1152/jappl.1987.63.3.1236
Р. и Кавана П.Р. (1987). Взаимосвязь между механикой бега на длинные дистанции, экономичностью бега и результативностью. J Appl Physiol . 63, 1236–1245. doi: 10.1152/jappl.1987.63.3.1236
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уильямс, К.Р., и Кавана, П.Р. (1986). «Биомеханические корреляции с экономичностью бега у элитных бегунов на длинные дистанции», в Proceedings of the North American Congress on Biomechanics (Montreal, QC), 287–288.
Академия Google
Почему четвероногие животные лучше бегают на короткие дистанции — ScienceDaily
Многие четвероногие млекопитающие могут развивать значительно более высокую скорость бега, чем двуногие люди.Животные, прекрасно приспособленные к бегу на короткие дистанции, такие как гепарды или антилопы, отличаются стройной формой тела, длинными ногами и особенно подвижным позвоночником, позволяющим достигать очень высоких скоростей при беге. Междисциплинарная группа, включающая исследователей из Института зоологии Кельнского университета, разработала модель, которая учитывает эти характеристики и может рассчитать максимальную скорость бега для животных любого размера. Результаты их исследования были опубликованы в статье «Правила беговой формулы природы: мышечная механика во время поздней стойки — ключ к объяснению максимальной скорости бега» в журнале Journal of Theoretical Biology .
Результаты их исследования были опубликованы в статье «Правила беговой формулы природы: мышечная механика во время поздней стойки — ключ к объяснению максимальной скорости бега» в журнале Journal of Theoretical Biology .
Олимпийские игры в Токио не за горами, и одним из самых ярких моментов станет мужской спринт на 100 метров. Лучшие спринтеры могут развивать скорость бега почти до 45 км/ч. Звучит впечатляюще, но на самом деле это не так по сравнению с бегом в животном мире. Это лишь примерно соответствует максимальной скорости домашней кошки. Гепарды могут бегать более чем в два раза быстрее (более 100 км/ч), но другие животные, такие как антилопы (90 км/ч) или даже бородавочники и зайцы (чуть менее 60 км/ч), обогнали бы людей-спринтеров.
Исследовательская группа под руководством доктора Михаэля Гюнтера (Университет Штутгарта) исследовала физические и биологические факторы, от которых зависит максимальная скорость этих животных. В процессе исследователи разработали исчерпывающие ответы на вопросы о важности конструкции тела, такие как: «Почему животные среднего размера достигают максимальной скорости в природе?», «Какие основные характеристики определяют максимальную скорость бега?» и «Какие характеристики ограничивают скорость?»
В основе их теоретической работы лежит физический баланс движущей силы ног и сопротивления воздуха, которое необходимо преодолеть, а также инерции движущих мышц. Они показывают своего рода основной путь изменения структурной формы тела животных в зависимости от размера тела (аллометрия) при адаптации к быстрому передвижению с помощью ног. «Этот основной путь описывает, как форма организма должна изменяться в зависимости от размера тела, чтобы достичь высокой скорости бега, и как конкретные формы влияют на максимальную скорость, которую можно достичь», — сказал доктор Том Вейхманн из Университета Нью-Йорка. Кёльнский институт зоологии.
Они показывают своего рода основной путь изменения структурной формы тела животных в зависимости от размера тела (аллометрия) при адаптации к быстрому передвижению с помощью ног. «Этот основной путь описывает, как форма организма должна изменяться в зависимости от размера тела, чтобы достичь высокой скорости бега, и как конкретные формы влияют на максимальную скорость, которую можно достичь», — сказал доктор Том Вейхманн из Университета Нью-Йорка. Кёльнский институт зоологии.
Классический пример — мышь и слон.Мышь размером со слона была бы просто нежизнеспособна, потому что ее кости сломались бы под собственным весом. У слонов гораздо более толстые и тяжелые кости относительно их веса, а также гораздо более длинные и прямые ноги. Эти особенности делают возможными огромные размеры животных. Однако тяжелые кости и прямые ноги ограничивают их максимальную скорость, которая намного ниже, чем у гепардов, хотя ноги слона намного длиннее.
Однако максимальная скорость зависит не только от размера, но и от конструкции, например от количества ног и подвижности позвоночника. Например, многие четвероногие млекопитающие могут развивать гораздо более высокую скорость бега, чем двуногие конструкции, такие как люди и птицы, потому что они могут галопировать, используя мышцы туловища для движения. «Однако, если животные становятся слишком тяжелыми, даже более мощные мышцы не помогут, потому что более крупным мышцам требуется больше времени, чтобы сокращаться на максимальной скорости. Соответственно, предел веса, выше которого скорость бега снова начинает снижаться, составляет около 50 кг, что довольно близко к среднему весу гепардов и вилорогов, самых быстрых спринтеров на нашей планете», — пояснил Вейхманн.
Например, многие четвероногие млекопитающие могут развивать гораздо более высокую скорость бега, чем двуногие конструкции, такие как люди и птицы, потому что они могут галопировать, используя мышцы туловища для движения. «Однако, если животные становятся слишком тяжелыми, даже более мощные мышцы не помогут, потому что более крупным мышцам требуется больше времени, чтобы сокращаться на максимальной скорости. Соответственно, предел веса, выше которого скорость бега снова начинает снижаться, составляет около 50 кг, что довольно близко к среднему весу гепардов и вилорогов, самых быстрых спринтеров на нашей планете», — пояснил Вейхманн.
Модель можно применить даже к фантастическим существам. Например, гигантский паук Шелоб из J.R.R. Толкиновский «Властелин колец» мог развивать максимальную скорость около 60 км/ч. С точки зрения геометрии человеческого тела модель показывает, что лучшие спринтеры в спорте уже очень близки к оптимальной скорости. Помимо технических приложений, таких как специальные кроссовки или экзоскелеты, обеспечивающие удлинение рычагов или дополнительную эластичность, только более длинные ноги или более эластичные сухожилия позволили бы развивать еще более высокие скорости.
Источник истории:
Материалы предоставлены Кёльнским университетом . Примечание. Содержимое можно редактировать по стилю и длине.
Что такое скорость процессора и почему это важно?
Благодаря технологиям, целям повышения производительности, более быстрому Интернету и большему количеству устройств мы создали потребность в скорости, где бы мы ни находились. Мы привыкли получать результаты мгновенно и ожидаем, что наши устройства будут соответствовать нашим запросам, когда мы многозадачны на протяжении всей жизни.Компьютерные процессоры и их тактовая частота — это две особенности, которые мы чаще всего связываем с высокопроизводительной и быстрой технологией.
Скорость процессора компьютера (скорость ЦП) является одним из наиболее важных элементов, которые следует учитывать при сравнении компьютеров. ЦП часто называют «мозгом» вашего компьютера, поэтому обеспечение его правильной работы очень важно для долговечности и функциональности вашего компьютера. Понимание того, что обеспечивает хорошую скорость процессора, начинается с понимания того, что именно делает процессор и что делают его компоненты для улучшения функциональности вашего компьютера.
Понимание того, что обеспечивает хорошую скорость процессора, начинается с понимания того, что именно делает процессор и что делают его компоненты для улучшения функциональности вашего компьютера.Давайте разберемся в том, что делает ваш процессор быстрым, количество ядер в сравнении с тактовой частотой, что делает их важными и на что обращать внимание при покупке нового компьютера.
Что такое процессор ПК и для чего он?
Центральный процессор или ЦП — это часть оборудования, которая позволяет вашему компьютеру взаимодействовать со всеми установленными приложениями и программами. Процессор интерпретирует инструкции программы и создает вывод, с которым вы взаимодействуете, когда используете компьютер.
Процессор состоит из оборудования, которое работает вместе для доставки информации, позволяя вашему компьютеру выполнять задачи, которые вы запрашиваете, когда открываете приложение или вносите изменения в файл. Независимо от того, обрабатывается он быстро или мучительно медленно, это может оказать большое влияние на ваш опыт работы с компьютером.
Ядра процессора и тактовая частота определяют, сколько информации может быть получено за раз и как быстро эта информация может быть обработана на вашем компьютере. Скорость, с которой ядра вашего компьютера и тактовая частота работают вместе, считается скоростью его обработки.
Ядра процессора и тактовая частота
Ядра процессора и тактовая частота — это очень разные функции, но они работают для достижения одной цели. Многие технические специалисты говорят о том, чему вы должны уделить больше внимания при покупке компьютера, но они в равной степени зависят друг от друга, чтобы помочь вашему компьютеру работать наилучшим образом.
Знание различий между ними может помочь вам лучше понять, что делает каждый из них и какая скорость процессора вам нужна в зависимости от того, как вы планируете использовать свой компьютер.Если вы планируете использовать свой компьютер для сложного редактирования видео, а не только для стандартных программ и просмотра веб-страниц, у вас будут другие требования к ядру процессора и тактовой частоте. Давайте рассмотрим эти две технологии и цифры, на которые следует обращать внимание при сравнении компьютеров.
Давайте рассмотрим эти две технологии и цифры, на которые следует обращать внимание при сравнении компьютеров.
Что такое ядро процессора?
Ядра процессора — это отдельные процессорные блоки в составе центрального процессора (ЦП) компьютера. Ядро процессора получает инструкции от одной вычислительной задачи, работая с тактовой частотой, чтобы быстро обрабатывать эту информацию и временно сохранять ее в оперативной памяти (ОЗУ).Постоянная информация сохраняется на вашем жестком диске, когда вы запрашиваете ее.Большинство компьютеров теперь имеют несколько ядер процессора, что позволяет вашему компьютеру выполнять несколько задач одновременно. Возможность запускать многочисленные программы и запрашивать несколько задач, таких как внесение изменений в документ, просмотр видео, открытие новой программы, стала возможной благодаря многоядерным процессорам.
Для сложных видеоигр или программ очень важно иметь ЦП, способный справляться с такой информацией, как быстро распространяемые аудио- и видеопотоки. В цифровую эпоху, когда мы все являемся экспертами в многозадачности, процессорные ядра становятся все более важными для пользователей компьютеров.
В цифровую эпоху, когда мы все являемся экспертами в многозадачности, процессорные ядра становятся все более важными для пользователей компьютеров.
Что такое тактовая частота?
Тактовая частота процессора компьютера определяет, насколько быстро центральный процессор (ЦП) может извлекать и интерпретировать инструкции.Это помогает вашему компьютеру выполнять больше задач, выполняя их быстрее.
Тактовые частоты измеряются в гигагерцах (ГГц), при этом большее число соответствует более высокой тактовой частоте. Многоядерные процессоры были разработаны, чтобы помочь процессорам работать быстрее, поскольку увеличить тактовую частоту стало сложнее. Более высокая тактовая частота означает, что вы увидите, что задачи, заказанные вашим ЦП, будут выполняться быстрее, что сделает вашу работу беспроблемной и сократит время, которое вы ожидаете, чтобы взаимодействовать с вашими любимыми приложениями и программами.
Как выбрать большее количество ядер процессора или более высокую тактовую частоту?
Как мы упоминали ранее, для работы вашего компьютера важны как ядра процессора, так и тактовая частота. Покупка компьютера с несколькими ядрами и сверхвысокой тактовой частотой звучит идеально, но что все это на самом деле означает для функциональности вашего компьютера?По сути, наличие высокой тактовой частоты, но всего одного или двух ядер означает, что ваш компьютер сможет быстро загружать одно приложение и взаимодействовать с ним.И наоборот, наличие большего количества ядер процессора, но более низкой тактовой частоты означает, что ваш компьютер может работать с большим количеством приложений одновременно, но каждое из них может работать немного медленнее.
При сравнении компьютеров важнее всего думать о своем образе жизни. Не всем нужен одинаковый уровень вычислительных скоростей или ядер. Мы немного обсудим, чем игровые компьютеры и повседневные рабочие или персональные компьютеры различаются, когда речь заходит об этих функциях. Во-первых, мы узнаем, что это означает для ноутбуков и настольных компьютеров.
Во-первых, мы узнаем, что это означает для ноутбуков и настольных компьютеров.
Какова хорошая скорость процессора для ноутбука по сравнению с настольным компьютером?
Процессоры ноутбуков отличаются от процессоров настольных компьютеров. Если вам интересно, что такое хорошая скорость процессора для ноутбука или настольного компьютера, или просто какой стиль лучше всего подходит для вас, прочитайте важные различия, которые следует учитывать, прежде чем делать какие-либо шаги по покупке.
Примечание. Процессоры также могут влиять на аппаратное обеспечение компьютера, поэтому их важно учитывать, если у вас есть особые требования к оборудованию, например портативность ноутбука, или вам нужна надежность настольного компьютера с двумя дисплеями.
Ноутбуки
Как правило, ноутбуки имеют меньшую мощность и гибкость, когда речь идет о процессорах. Они, очевидно, очень удобны для пользователей, которым нравится мобильность ноутбука, но если вам нужен сверхскоростной процессор или высокая тактовая частота, вы можете рассмотреть настольный компьютер для удовлетворения ваших потребностей в обработке.
Благодаря удивительным разработкам в области многоядерных процессоров и методов гиперпоточности ноутбуки теперь могут выстоять.Большинство ноутбуков оснащены двухъядерными процессорами, которые удовлетворяют потребности большинства повседневных пользователей. А некоторые используют четырехъядерные процессоры, которые могут повысить вычислительные возможности вашего ноутбука.
Настольные компьютеры
Настольные компьютеры могут генерировать больше энергии, чем ноутбуки, благодаря их надежному оборудованию, которое обеспечивает большую вычислительную мощность и более высокую тактовую частоту. Поскольку у них больше места в корпусе, чем у ноутбука, настольные компьютеры обычно имеют более совершенные системы охлаждения, что позволяет процессору продолжать усердно работать, не перегреваясь.
ЦП настольного компьютера обычно можно снять, в отличие от ЦП ноутбука, который встроен в материнскую плату. Это означает, что ЦП легче модернизировать или заменить на настольном ПК, чем на ноутбуке. Если вы выберете правильную для вас частоту процессора, вам не придется возиться с вашим процессором.
Если вы выберете правильную для вас частоту процессора, вам не придется возиться с вашим процессором.
Независимо от того, используете ли вы ноутбук или настольный компьютер, в конечном итоге вы захотите подумать о том, для чего вы планируете использовать свой компьютер, поскольку это напрямую связано с необходимой вам скоростью процессора компьютера.
Жажда скорости
Игровые процессоры
Игры с годами становятся все более сложными и, кажется, совершенствуются с каждым днем. Все эти дополнительные функции и реалистичные впечатления требуют процессора, способного поддерживать вашу игру. В большинстве игр используется от 1 до 4 ядер, а многим требуется даже больше процессорных ядер для оптимальной работы. Четырехъядерный процессор занимает безопасную зону, когда речь заходит о ядерных блоках.
Такие игры, как World of Warcraft , постоянно улучшают игровой процесс благодаря обновленным игровым возможностям и игровым сценариям, требующим более мощной обработки. В играх с интенсивным использованием ЦП используется многоядерная технология, позволяющая объединить графику, звук и игру для создания гиперреалистичного игрового процесса.
В играх с интенсивным использованием ЦП используется многоядерная технология, позволяющая объединить графику, звук и игру для создания гиперреалистичного игрового процесса.Одноядерный процессор лучше всего подходит для выполнения одиночных задач, но это может повлиять на вашу игру и замедлить работу. Больше ядер может помочь достичь более высокого качества игрового процесса.
Если вы серьезный геймер, который ценит целостность разработанного разработчиком опыта, вы можете рассмотреть четырехъядерный или более мощный процессор, такой как процессор Intel® Core™ i7-8750H, установленный в 15-дюймовом игровом ноутбуке HP OMEN. .Этот мощный процессор использует 6 ядер для отображения игрового пространства и реагирования на игровую технику с беспрецедентной гибкостью. Тактовая частота от 3,5 ГГц до 4,0 ГГц обычно считается хорошей тактовой частотой для игр, но важнее иметь хорошую однопоточную производительность. Это означает, что ваш ЦП хорошо справляется с пониманием и выполнением отдельных задач.
Не путать с одноядерным процессором. Наличие большего количества ядер означает, что ваш ЦП может понимать инструкции нескольких задач, а оптимальная однопоточность означает, что он может обрабатывать каждую из них по отдельности и очень хорошо.
Видеоигры переносят вас в другой мир и дают вам возможность исследовать новые территории. Не позволяйте недостатку вычислительной мощности лишить магии ваш мир.
Процессоры для повседневного использования
Двухъядерный процессор обычно лучше всего подходит для повседневного использования. Он может работать в многозадачном режиме и сокращать время, затрачиваемое на ожидание открытия приложений или обновлений. Четырехъядерный процессор может помочь вам поднять производительность на новый уровень и обеспечить согласованность для лучшего опыта работы с компьютером, независимо от того, над чем вы работаете.
Если вы более творческий человек, который ежедневно редактирует видео или запускает сложные приложения, вы можете подумать о приобретении компьютера с большим количеством процессорных ядер и более высокой тактовой частотой, чтобы ваши приложения могли работать без сбоев. 15-дюймовая мобильная рабочая станция HP ZBook оснащена 6-ядерным процессором, разработанным для интенсивного редактирования и дизайна для творческих людей. потоковая передача и проверка электронной почты.Возможно, вам нужен ноутбук HP 14z с двухъядерным процессором для ежедневного базового использования. Эта модель способна легко справляться с общими задачами в традиционном корпусе, который прост в использовании.
15-дюймовая мобильная рабочая станция HP ZBook оснащена 6-ядерным процессором, разработанным для интенсивного редактирования и дизайна для творческих людей. потоковая передача и проверка электронной почты.Возможно, вам нужен ноутбук HP 14z с двухъядерным процессором для ежедневного базового использования. Эта модель способна легко справляться с общими задачами в традиционном корпусе, который прост в использовании.Высокопроизводительные вычислительные процессоры
Под высокопроизводительными вычислениями понимается использование компьютера, включающее чрезвычайно сложные программы, требующие большого объема данных. Высокопроизводительные пользователи часто являются инженерами, исследователями, а также военными или государственными пользователями.
Эти пользователи последовательно запускают несколько программ и постоянно извлекают и вводят информацию в программные системы.Этот тип вычислений обычно требует более продвинутого процессора и более высокой тактовой частоты, чтобы не отставать.
Процессоры для иммерсивных вычислений и виртуальной реальности (VR)
Подобно играм, дополненная реальность и виртуальная реальность зависят от высококачественной графики, звука и навигационных функций. Чтобы действительно почувствовать себя в новой реальности, необходимо иметь многоядерный процессор с высокой тактовой частотой.Выберите компьютер, который подходит именно вам
Большинство людей знают, как выглядит их использование компьютера; либо вы геймер, либо нет, вы используете свой компьютер каждый день или нет.Знание этой информации о собственных привычках облегчает выбор процессора.
Если вы одновременно запускаете много приложений или играете в сложные игры, вам, скорее всего, понадобится 4-х или даже 8-ядерный процессор. Если вы просто ищете компьютер для эффективного выполнения основных задач, двухъядерный процессор, вероятно, подойдет для ваших нужд.
Для вычислений с интенсивным использованием ЦП, таких как редактирование видео или игры, вам потребуется более высокая тактовая частота, близкая к 4,0 ГГц, в то время как базовые вычислительные потребности не требуют такой повышенной тактовой частоты.
Об авторе
Софи Сируа (Sophie Sirois) является автором статьи для HP® Tech Takes. Софи — специалист по созданию контента из Сан-Диего, освещающий последние новости в области технологий и цифровых технологий.Пошаговые вариации человеческого бега показывают, как люди бегают, не падая
Основные версии:
1A) Авторы склонны преувеличивать новизну своих открытий, предполагая, что предыдущая работа, основанная на модели бега с пассивной пружинной массой, не может объяснить стабильный бег.Предыдущая работа, основанная на механике бега с пружинной массой, была разработана, чтобы показать, как пассивно-эластичные свойства могут уменьшить работу мышц, необходимых для управления движением ног и тела.
 Но эти прошлые исследования не утверждают, что мышечная работа сама по себе не нужна. Как отмечалось в собственной работе автора (Srinivasan, 2011) и более ранних работах (Alexander, 1997; Ruina et al., 2005) (в дополнение к другим — например, Biewener and Daley, 2007), пружинящее поведение ног может быть результатом неупругая конечность, в которой выполняется отрицательная работа мышц, за которой следует равная положительная работа.Авторам следует пересмотреть, чтобы более точно отразить это понимание и подход прошлых специалистов по динамике бега по отношению к их собственной работе. Мы понимаем, что некоторые из этих более ранних исследований, возможно, преувеличивали их влияние на динамическую стабильность во время бега. Тем не менее, мы призываем авторов сосредоточиться на своих новых данных и модели и позволить этому говорить о вкладе их исследования. Примеры перечислены ниже:
Но эти прошлые исследования не утверждают, что мышечная работа сама по себе не нужна. Как отмечалось в собственной работе автора (Srinivasan, 2011) и более ранних работах (Alexander, 1997; Ruina et al., 2005) (в дополнение к другим — например, Biewener and Daley, 2007), пружинящее поведение ног может быть результатом неупругая конечность, в которой выполняется отрицательная работа мышц, за которой следует равная положительная работа.Авторам следует пересмотреть, чтобы более точно отразить это понимание и подход прошлых специалистов по динамике бега по отношению к их собственной работе. Мы понимаем, что некоторые из этих более ранних исследований, возможно, преувеличивали их влияние на динамическую стабильность во время бега. Тем не менее, мы призываем авторов сосредоточиться на своих новых данных и модели и позволить этому говорить о вкладе их исследования. Примеры перечислены ниже: Спасибо за конструктивные замечания.Мы согласны с замечаниями рецензентов о том, что многие авторы, в том числе те, кто использовал парадигму модели пружинной массы и впервые разработал ее, осознали ограничения и упрощения моделей пружинной массы на протяжении многих лет, в то время как, возможно, некоторые авторы восприняли метафору слишком далеко. рецензенты соглашаются. Мы также согласны с тем, что первоначально модели с пружинной массой были вдохновлены тем, как пассивные эластичные свойства могут помочь уменьшить мышечную работу, необходимую для бега, и были полезны для решения этих задач.Поэтому, как предполагают рецензенты, мы больше сосредоточились на наших результатах, а не на предыдущих моделях, подобных пружинной массе. В частности, мы внесли следующие изменения, чтобы уменьшить или удалить нашу критику моделей, подобных пружинной массе, и/или более явно приписать ее слабость предыдущим авторам (что мы, возможно, сделали более неявно в предыдущей версии через цитаты в скобках). Ниже мы перечислили многие из этих правок в дополнение к дополнительным, перечисленным в ответ на комментарии 1B, 1C, 1D и 1E.
рецензенты соглашаются. Мы также согласны с тем, что первоначально модели с пружинной массой были вдохновлены тем, как пассивные эластичные свойства могут помочь уменьшить мышечную работу, необходимую для бега, и были полезны для решения этих задач.Поэтому, как предполагают рецензенты, мы больше сосредоточились на наших результатах, а не на предыдущих моделях, подобных пружинной массе. В частности, мы внесли следующие изменения, чтобы уменьшить или удалить нашу критику моделей, подобных пружинной массе, и/или более явно приписать ее слабость предыдущим авторам (что мы, возможно, сделали более неявно в предыдущей версии через цитаты в скобках). Ниже мы перечислили многие из этих правок в дополнение к дополнительным, перечисленным в ответ на комментарии 1B, 1C, 1D и 1E.
Оригинал: «Классическая парадигма моделирования управления бегом предполагает, что человеческая нога ведет себя как линейная пружина (Бликхан, 1989; МакМахон и Ченг, 1990; Бликхан и Фулл, 1993)».
Пересмотрено: «Одна из классических парадигм моделирования управления бегом предполагает, что человеческая нога ведет себя как линейная пружина (Бликхан, 1989; МакМэхон и Ченг, 1990; Бликхан и Фулл, 1993). Эта парадигма использовалась для аргументации того, как пассивно-эластические свойства могут уменьшить мышечную работу, необходимую для передвижения (Александер и Вернон, 1975; Александр, 1990), и была полезна при изучении различных аспектов передвижения в упрощенном виде.
Эта парадигма использовалась для аргументации того, как пассивно-эластические свойства могут уменьшить мышечную работу, необходимую для передвижения (Александер и Вернон, 1975; Александр, 1990), и была полезна при изучении различных аспектов передвижения в упрощенном виде.
Оригинал: «Эти модели обычно основаны на подгонке к среднему периодическому движению центра масс во время 38 прогонов (Бликхан и Фулл, 1993; Гейер и др., 2006; Шринивасан и Холмс, 2008). Однако понимание стабильности бега требует понимания того, как контролируются отклонения от среднего движения 40».
Пересмотрено: «Эти модели успешно соответствуют среднему периодическому движению центра масс во время бега (Бликхан и Фулл, 1993; Гейер и др., 2006; Шринивасан и Холмс, 2008).Понимание стабильности бега требует понимания того, как контролируются отклонения от среднего движения».
Оригинал: «Пружинная механика ног не может объяснить, как контролируются отклонения от среднего движения (Maus et al. , 2015)».
, 2015)».
Пересмотрено: «Ранее было признано, что пружинящая механика ног не может объяснить, как отклонения от среднего движения контролируются и в конечном итоге ослабляются (например, Ghigliazza et al., 2005; Biewener and Daley, 2007; Maus et al., 2015)».
Оригинал: «Здесь мы используем более общие модели передвижения человека, основанные на ньютоновской механике, для реализации управления бегом, не делая никаких параметрических предположений, подобных пружинам (Srinivasan 2011)».
Пересмотрено: «Здесь мы исследуем роль активного мышечного контроля в стабильности бега, используя более общие модели передвижения человека, основанные на ньютоновской механике (Srinivasan, 2011)».
Отредактированная версия: В следующем предложении мы удалили ссылку на модель пружинной массы.
«Мы раскрываем, как достигается такой центр управления массами. Затем мы реализуем этот созданный человеком контроллер на простой математической модели двуногого человека (Srinivasan, 2011), показывая, что эта модель двуногого человека работает без падений, несмотря на непрекращающиеся шумоподобные возмущения, большие внешние возмущения и на неровной местности».
Пересмотрено: «Здесь мы использовали более простую «непараметрическую» модель для непосредственного описания управления усилием или активацией опорной ноги (рис. 8)».
1B) Большинство тех, кто моделировал устойчивый бег как систему пружины и массы с пружинной динамикой ног, не предполагают, что эта пассивная модель является точным представлением того, как управляется бег, или что она обязательно приведет к устойчивым периодическим движение после возмущения.Телескопическая модель ноги, контролируемая мышцами одного коленного сустава, которая показывает чистую работу ноги (как отрицательную, так и положительную), в этом отношении неудивительна и является ожидаемым результатом. Пожалуйста, подумайте лучше о том, что исследователи прошлого, которые анализируют бег как динамику пружины и массы, явно или неявно признают, что необходима некоторая мышечная работа, чтобы контролировать изменяющиеся во времени движения состояния ЦМ бегуна для стабильного периодического движения.

Спасибо за эти замечания. В предыдущем написании абзаца мы недостаточно ясно выразились в том, что приписываем некоторые из этих рассуждений предшествующим исследователям, хотя и цитировали их в скобках.Мы переписали соответствующий абзац, чтобы лучше отразить это предварительное понимание:
«Ожидается, что любой работающий контроллер, который достигает асимптотической устойчивости, должен будет выполнять результирующую механическую работу в ответ на возмущения, которые уменьшают или увеличивают механическую энергию тела (Ghigliazza et al., 2005; Srinivasan and Holmes, 2008; Maus et al. , 2015). Наши результаты согласуются с такими ожиданиями, как показано рабочими циклами с чистой механической работой на рисунке 11.Энергосберегающее пружинистое поведение ноги не допускает такой чистой механической работы и может обеспечить лишь частичную асимптотическую устойчивость, не имея возможности справляться с возмущениями, изменяющими энергию (как отмечено Ghigliazza et al. (2005)).
1C) Введение «Здесь, напротив, мы избегаем допущений, подобных пружинной массе, и характеризуем управление с точки зрения того, как люди модулируют величину и направление силы своей ноги». Это несколько вводит в заблуждение, поскольку предположения о пружинной массе, сделанные ранее другими рабочими, должны были исследовать, в какой степени бег (рысь, прыжки) можно объяснить пассивной динамикой, признавая при этом, что некоторая работа мышц для стабилизации бега (или для перехода подъем/спуск, изменение направления) необходимо.Большинство предшествующих работников не заявляют, что бег является чисто пассивным и не требует мышечной работы для контроля и устойчивости. Пожалуйста, более точно отразите понимание и работу в этой области здесь.
Спасибо за эти замечания. Теперь мы отредактировали следующее предложение:
«Здесь, напротив, мы избегаем допущений, подобных пружинной массе, и характеризуем контроль с точки зрения того, как люди модулируют величину и направление силы своей ноги». до «Здесь мы напрямую характеризуем контроль с точки зрения того, как люди модулируют величину и направление силы своих ног во время бега.
до «Здесь мы напрямую характеризуем контроль с точки зрения того, как люди модулируют величину и направление силы своих ног во время бега.
1D) Подраздел «Человеческий контроллер стабилизирует минимальную модель двуногого бега»: «Стабильный бег не может быть чисто пассивным и предполагает активную работу ног». Пожалуйста, исправьте, чтобы не преувеличивать значимость моделирования и выводов авторов.
Спасибо. Как и было предложено, мы отредактировали первоначальный подраздел «Стабильный бег не может быть чисто пассивным и включает в себя активную работу ног», чтобы он был более «иллюстративным» в отношении предыдущих ожиданий, более явно признавая различных авторов по этой теме.
«Ненулевая работа ноги для изменяющих энергию возмущений.
Рисунок 11 иллюстрирует рабочий цикл ноги для невозмущенного пробега (чистая нулевая работа) и когда положительные возмущения применяются к боковым и продольным скоростям, а также к вертикальным положениям. Все такие положительные возмущения приводят к чистой отрицательной работе на первом шаге после возмущения, что отражается в рабочих циклах с чистой отрицательной площадью внутри них. Такая чистая положительная или отрицательная работа ног явно необходима для восстановления после возмущений, которые изменяют общую механическую энергию бегуна, как это было признано в предшествующих обсуждениях модели бега с нейтральной энергией пружины и массы (Ghigliazza et al., 2005; Бивенер и Дейли, 2007 г.; Шринивасан и Холмс, 2008 г.)».
Все такие положительные возмущения приводят к чистой отрицательной работе на первом шаге после возмущения, что отражается в рабочих циклах с чистой отрицательной площадью внутри них. Такая чистая положительная или отрицательная работа ног явно необходима для восстановления после возмущений, которые изменяют общую механическую энергию бегуна, как это было признано в предшествующих обсуждениях модели бега с нейтральной энергией пружины и массы (Ghigliazza et al., 2005; Бивенер и Дейли, 2007 г.; Шринивасан и Холмс, 2008 г.)».
1E) Раздел для обсуждения: Псевдоупругое поведение ноги бегуна, которое на самом деле может быть полностью основано на срабатывании (отрицательная работа, за которой следует эквивалентная положительная работа) – как показывают ссылки на Ruina et al., 2005 и Alexander, 1997, а также собственные прошлые работы автора (Srinivasan, 2011).
Спасибо за эти замечания. Теперь мы удалили предложение, которое ранее было «Наши результаты предполагают…», как описано в ответе на комментарий рецензента 1B.
Что касается седьмого абзаца раздела «Обсуждение», мы предполагаем, что рецензенты соглашаются с нами и просто обращают наше внимание на наш абзац, касающийся предыдущих обсуждений псевдоэластичного поведения ног при беге. Мы оставили этот абзац в основном нетронутым, немного отредактировав его следующим образом:
«Действительно, обычно считается, что даже стационарное движение центра масс, похожее на пружину, при беге происходит из-за значительного мышечного действия и называется псевдоупругим (Ruina et al., 2005) или псевдосовместимые (Александр, 1997). Примечательно, что оптимальные с точки зрения энергии беговые движения в моделях без подпружинивания ног создают сходные траектории центра масс, подобные пружине (Srinivasan, 2011), при этом мышцы ног выполняют одинаковое количество положительной и отрицательной работы».
2) Существует большое количество литературы, посвященной изучению вариабельности походки (например, GEM Дингвелла, UCM Чанга, корреляции долгосрочных шумов Хаусдорфа, анализ основных компонентов Лакуанити и т.
 д.).). Эти подходы в основном основаны на характеристике изменчивости кинематических показателей как способе оценки «стабильности», но они не могут много сказать о лежащем в основе контроле, и это ограничило их влияние на более фундаментальное понимание контроля механики походки. Представленный здесь подход является давно назревшей, более формальной структурой идентификации линейных систем, основанной на механике Ньютона-Эйлера. Возможно, стоит признать это и сравнить/сопоставить полезность этих двух подходов (например,г., диагноз против предсказания) без чрезмерной критики предыдущих подходов. Например. сосредоточив одно или несколько предложений в обсуждении на ключевых новых способностях, которые дополняют предыдущие подходы, и на том, как их можно использовать вместе в будущих исследованиях.
д.).). Эти подходы в основном основаны на характеристике изменчивости кинематических показателей как способе оценки «стабильности», но они не могут много сказать о лежащем в основе контроле, и это ограничило их влияние на более фундаментальное понимание контроля механики походки. Представленный здесь подход является давно назревшей, более формальной структурой идентификации линейных систем, основанной на механике Ньютона-Эйлера. Возможно, стоит признать это и сравнить/сопоставить полезность этих двух подходов (например,г., диагноз против предсказания) без чрезмерной критики предыдущих подходов. Например. сосредоточив одно или несколько предложений в обсуждении на ключевых новых способностях, которые дополняют предыдущие подходы, и на том, как их можно использовать вместе в будущих исследованиях.Спасибо за эти замечания. Теперь мы добавили новый абзац, который показывает, как наши методы могут дополнять эти другие методы:
«Некоторые прошлые работы по выводу устойчивости из изменчивости были сосредоточены на кинематических показателях изменчивости, таких как множители Флоке, показатели Ляпунова с конечным временем (Dingwell et al. , 2001) и долгосрочные корреляции вариабельности ходьбы и бега (Hausdorff et al., 1995; Jordan et al., 2006; Kaipust et al., 2012). Такие меры могут обеспечить дискриминационные диагностические меры (Kaipust et al., 2012), но не пытаются предоставить причинно-следственную связь о том, как контролируется передвижение. Наш подход здесь, основанный на механике Ньютона-Эйлера, способен обнаружить потенциальные причинно-следственные стратегии, лежащие в основе стабильности движения, и, в более широком смысле, может помочь в лечении патологических нестабильных движений в дополнение к диагностике.В других прошлых исследованиях использовались варианты анализа главных компонентов (Cappellini et al., 2006; Maus et al., 2015), чтобы продемонстрировать, что внутренняя изменчивость человеческого передвижения может находиться в многообразии более низкого измерения (Cappellini et al., 2006; Maus et al., 2015; Dingwell et al., 2010; Chang et al., 2009; Yen et al., 2009; Maus et al., 2015). Здесь мы использовали уменьшение размерности, основанное на физике, чтобы изучить доминирующие стратегии управления, сосредоточив внимание на том, как центр масс контролируется силами.
, 2001) и долгосрочные корреляции вариабельности ходьбы и бега (Hausdorff et al., 1995; Jordan et al., 2006; Kaipust et al., 2012). Такие меры могут обеспечить дискриминационные диагностические меры (Kaipust et al., 2012), но не пытаются предоставить причинно-следственную связь о том, как контролируется передвижение. Наш подход здесь, основанный на механике Ньютона-Эйлера, способен обнаружить потенциальные причинно-следственные стратегии, лежащие в основе стабильности движения, и, в более широком смысле, может помочь в лечении патологических нестабильных движений в дополнение к диагностике.В других прошлых исследованиях использовались варианты анализа главных компонентов (Cappellini et al., 2006; Maus et al., 2015), чтобы продемонстрировать, что внутренняя изменчивость человеческого передвижения может находиться в многообразии более низкого измерения (Cappellini et al., 2006; Maus et al., 2015; Dingwell et al., 2010; Chang et al., 2009; Yen et al., 2009; Maus et al., 2015). Здесь мы использовали уменьшение размерности, основанное на физике, чтобы изучить доминирующие стратегии управления, сосредоточив внимание на том, как центр масс контролируется силами.
3) Принятый здесь подход к моделированию оформлен в виде линейного идентификатора системы, который подходит для небольших возмущений, в данном случае пошагового изменения среднего поведения. Рисунок 8 начинает решать эту проблему в моделировании, но насколько хорошо этот подход применим к более явным, более крупным возмущениям на разных фазах походки, когда нелинейность может стать более важной? Каков план борьбы с этим? Пожалуйста, уточните это или лежащие в основе ограничения в обсуждении.
Спасибо за эти замечания. Теперь мы добавили следующие примечания в раздел «Обсуждение»:
«Мы сосредоточились на линейных отношениях между отклонениями состояния и контролем, поскольку они естественным образом подходят для небольших отклонений и возмущений, содержащихся в наших данных. В будущей работе мы надеемся изучить диапазон размеров возмущений, для которых это линейное описание является точным, путем сравнения этого линейного управления с ответами на эксперименты с большими возмущениями, а также сделать вывод о нелинейных описаниях, если они улучшат прогностическую способность.
4) Пожалуйста, обсудите потенциальные физиологические механизмы, лежащие в основе извлеченного контроллера, и предложите эксперименты, которые можно было бы провести для их выяснения, если у вас есть новый взгляд на это. Например, авторы упоминают комбинацию процессов прямой и обратной связи, но не особо подробно рассказывают о том, какой вклад могут вносить зрение, вестибулярный аппарат, проприоцепция и внутренняя модель мозжечка.
Спасибо за эти замечания. Теперь мы добавили следующий абзац в раздел «Обсуждение», касающийся сенсорной интеграции, которая может потребоваться для осуществления контроля с обратной связью:
. «Информация о состоянии центра масс или другая информация о состоянии тела, необходимая для контроля с обратной связью, может быть оценена нервной системой с помощью интегрированных сенсорных сигналов от зрения (Patia, 1997), проприоцептивных сенсоров (особенно, когда ступня находится на земле (Sainburg et al. , 1995)) и вестибулярные датчики (Angelaki and Cullen, 2008), возможно, в сочетании с прогнозирующими внутренними моделями (Walpert et al., 1995; Cullen, 2004). В будущей работе повторение приведенных здесь расчетов (например, рис. 5) для экспериментов, которые систематически блокируют или ухудшают (скажем, путем добавления сенсорного носа) один или несколько из этих датчиков, вполне может рассказать нам о вкладе этих датчиков в управление бегом. Мы предполагаем, что используется наиболее доступная релевантная сенсорная информация, возможно, аналогичная оценщику оптимального состояния (Kuo, 2005; Srinivasan, 2009), и ухудшение качества одного сенсора может привести к сенсорному перевзвешиванию в медленном временном масштабе (Carver et al., 2006; Аслендер и Петерка, 2014 г.)».
, 1995)) и вестибулярные датчики (Angelaki and Cullen, 2008), возможно, в сочетании с прогнозирующими внутренними моделями (Walpert et al., 1995; Cullen, 2004). В будущей работе повторение приведенных здесь расчетов (например, рис. 5) для экспериментов, которые систематически блокируют или ухудшают (скажем, путем добавления сенсорного носа) один или несколько из этих датчиков, вполне может рассказать нам о вкладе этих датчиков в управление бегом. Мы предполагаем, что используется наиболее доступная релевантная сенсорная информация, возможно, аналогичная оценщику оптимального состояния (Kuo, 2005; Srinivasan, 2009), и ухудшение качества одного сенсора может привести к сенсорному перевзвешиванию в медленном временном масштабе (Carver et al., 2006; Аслендер и Петерка, 2014 г.)».
5) Можно ли применить этот подход и к меньшим наборам данных от отдельных лиц? Насколько вариативен контроллер для разных людей и сколько данных необходимо для получения хорошей модели?
Спасибо за этот вопрос. Теперь мы добавили совершенно новый иллюстративный рисунок (рис. 12), который дает информацию по этим вопросам. Мы также добавили следующий абзац в раздел «Обсуждение», в котором упоминаются эти вопросы.
Теперь мы добавили совершенно новый иллюстративный рисунок (рис. 12), который дает информацию по этим вопросам. Мы также добавили следующий абзац в раздел «Обсуждение», в котором упоминаются эти вопросы.
«Результаты, которые мы представили, относятся к объединенным данным по всем субъектам. Выполнение регрессий для данных по отдельным субъектам показывает, что доминирующие условия в предполагаемых регуляторах одинаковы для всех субъектов; индивидуальная изменчивость расчетных результатов контроля показана на рис. 12а. На рис. 12b показано, как точность оцениваемого прироста контроля зависит от количества шагов, используемых для регрессии. Ожидается, что для таких линейных регрессий оценка ошибки (стандартное отклонение) уменьшится на N шаг , как 1∕ N 2 шаг , так что уменьшение ошибки в 10 раз требует 100-кратного уменьшения ошибки. увеличение размера выборки (Wang and Srinivasan, 2012; Hamilton, 1994).Эта зависимость от 90 133 N 90 134 90 189 шага 90 190 может помочь при выборе размеров выборки для будущих экспериментальных планов».
6) Введение «Многие работающие роботы продемонстрировали стабильную периодическую работу с использованием различных схем управления (Raibert, 1986; Chevallereau et al., 2005; Tajima et al., 2009)». Если роботы двигаются стабильно и периодически, как указано здесь, почему тогда бег людей и других животных не является стабильным и периодическим, как утверждает автор (Введение)? Предыдущее утверждение сделано, чтобы доказать важность мышечного срабатывания для контроля, чтобы достичь действительно стабильного периодического движения.Ясно, что и животным, и роботам необходимо выполнять некоторую мышечную работу, чтобы управлять движением. Это противоречие в формулировках необходимо устранить.
Спасибо, что указали на несоответствие в формулировках. Теперь мы отредактировали следующим образом, что, надеюсь, устраняет несоответствие.
«Некоторые бегущие роботы продемонстрировали стабильный бег (т., 2019)»
То есть мы отредактировали предложение, (1) удалив слово «периодический» применительно к работе робота и (2) также уточнив, что мы подразумеваем под «стабильной работой робота», то есть «бег без падения». Под устойчивым бегом мы подразумеваем не идеально периодическое движение, а именно то, что движение не приводит к падению. Мы надеемся, что наши правки устранили кажущееся несоответствие между этим предложением и предыдущим предложением о том, что человеческий бег не является идеально периодическим.
Под устойчивым бегом мы подразумеваем не идеально периодическое движение, а именно то, что движение не приводит к падению. Мы надеемся, что наши правки устранили кажущееся несоответствие между этим предложением и предыдущим предложением о том, что человеческий бег не является идеально периодическим.
7) Должны быть показаны регрессии для уравнений 1 и 2 (хотя бы в дополнительных материалах).
Спасибо за это предложение. Теперь мы включили в статью новый рисунок 3, на котором показана эта регрессия, приведенный ниже с заголовком.
”цикла шага, в частности, доли времени Ø стойки стойки (рис. 2b). Чтобы объяснить эту зависящую от фазы изменчивость силы за один шаг, мы вычисляем зависимую от фазы чувствительность ( F x , Fy, F z ) к состоянию центра масс следующим образом.Для каждого результата, скажем, F x , мы делим длительность стойки на 20 фаз и вычисляем линейную модель для F x на каждой из этих фаз, все с ( ẋ a , ẏ a , z и ) в качестве входов. Мы ссылаемся на коэффициенты в этих линейных моделях как функцию фазы Ø позиции как на зависящую от фазы чувствительность GRF (рис. 5) к соответствующей предикторной переменной в ( ẋ a , ẏ a , z и ).
Мы ссылаемся на коэффициенты в этих линейных моделях как функцию фазы Ø позиции как на зависящую от фазы чувствительность GRF (рис. 5) к соответствующей предикторной переменной в ( ẋ a , ẏ a , z и ).
8) Подраздел «Управление импульсами достигается за счет фазозависимой силовой модуляции»: «рассчитать фазозависимую чувствительность GRF к…» Как именно вычисляются эти фазозависимые чувствительности? (Обозначение оси Y для них необходимо определить/объяснить более четко.) Это частично объясняется (подраздел «Линейная регрессия между выходными данными и входными данными»), но может быть сделано более четким и явным при отображении и ссылке на эти паттерны на рисунке 4.
9) Рис. 5: Предсказуемость репозиции стопы при махе и состояние ЦМ при определении/расчете положения стопы – необходимо прояснить, как они определяются/рассчитываются.Результаты заявлены, но их основа не показана.
Спасибо за эти замечания. Теперь мы добавили следующий дополнительный абзац в раздел «Материалы и методы».
Теперь мы добавили следующий дополнительный абзац в раздел «Материалы и методы».
«В дополнение к регрессиям, описанным выше, с использованием состояния вершины полета в качестве предиктора, мы использовали состояние центра масс (Δ ẋ a , Δ ẏ a , Δ z a ) на разных фазах по сравнению с предыдущим шагом, чтобы предсказать каждый из выходных данных фазы стойки.В частности, для каждого результата фазы опоры мы выполнили 20 регрессий, каждая из которых использовала состояние центра масс в одной из 20 равноотстоящих фаз ходьбы по сравнению с предыдущим шагом, где один полный шаг определяется как начинающийся и заканчивающийся приземлением. Этот анализ позволяет исследовать предсказательную способность состояния центра масс на разных фазах. Для этих зависимых от фазы регрессий, в дополнение к использованию состояния центра масс в качестве предиктора, мы повторили расчеты, используя состояние маховой ноги (положение и скорость относительно центра масс), чтобы сравнить различные прогностические способности как на рисунке 6.
Мы также расширили подпись к рисунку с
.» Рис. 6. Управление махом ногой перед размещением ступни. Доля расстановки ступни в сторону (рисунок a) и расстановка ступни вперед-назад (рисунок b) дисперсии в начале стойки, прогнозируемая по состоянию центра масс (CoM) или состояние маховой ноги во время предыдущего шага (полет и стойка). Чтобы получить эту фигуру, была построена последовательность линейных моделей для прогнозирования положения стопы на основе состояния центра масс или состояния маховой стопы на разных этапах предыдущего шага.Мы наносим на график значение R 2 , соответствующее этим линейным моделям (то есть объясненную долю дисперсии), как функцию фазы походки, используемой для прогнозирования; фаза походки представлена как часть шага, начинающегося с начала предыдущей фазы опоры. Сплошная и пунктирная линии обозначают положение правой и левой стопы соответственно».
10) Рисунок 6: Отношения сила-скорость и сила-длина, показанные и реализованные в авторской модели, могут быть слишком простыми.
 F-V представляет собой обратную гиперболическую нелинейную зависимость, и удлиняющая сторона F-V гораздо более асимметрична, чем в модели авторов. (См. также Zajac, 1989). Как более реалистичная зависимость F-V (обратная гипербола для укорочения и более крутое и быстрое выравнивание силы по отношению к удлинению) повлияет на модель авторов?
F-V представляет собой обратную гиперболическую нелинейную зависимость, и удлиняющая сторона F-V гораздо более асимметрична, чем в модели авторов. (См. также Zajac, 1989). Как более реалистичная зависимость F-V (обратная гипербола для укорочения и более крутое и быстрое выравнивание силы по отношению к удлинению) повлияет на модель авторов?Спасибо за эти замечания. Теперь мы добавили следующие предложения, чтобы прокомментировать упрощение этой модели:
«Кроме того, мы сделали упрощающие предположения об архитектуре мышц, свойствах мышц (линейные отношения сила-скорость) и активации мышц, которые предназначены для захвата первичных качественных динамических характеристик мышц, а не для их точного количественного моделирования.Например, линейного соотношения сила-скорость может быть достаточно для создания демпфирующего поведения мышц при активации, но это демпфирующее поведение может усиливаться нелинейным соотношением сила-скорость. Будущая работа также будет включать в себя получение контроллеров для более сложных двуногих моделей и моделей мышц».